 ARTÍCULO ORIGINAL
ARTÍCULO ORIGINALARTÍCULO ORIGINAL
Estructura diamétrica y por estado, mortalidad y reclutamiento de Juglans jamaicensis C. DC., en el Parque Nacional Turquino
Diametric and status structure, mortality and recruitment of Juglans jamaicensis C. DC., in the Turquino National Park
Revista Cubana de Ciencias Forestales
Año 2015, Volumen 3, número 1
José Luis Rodríguez Sosa1, Calixto Aguilar Espinosa2, José Yulier Rodríguez Milanés3
1Máster en Ciencias Forestales. Departamento de Ingeniería Forestal, Universidad de Granma. Carretera a Manzanillo Km 17 ½. Peralejo, Bayamo. Granma. Cuba. teléf.: 0123-452249. Correo electrónico: jrodriguezs@udg.co.cu
RESUMEN
El trabajo se realizó en el Parque Nacional Turquino con el objetivo de analizar la estructura diamétrica, la mortalidad y el reclutamiento de Juglans jamaicensis C. DC., para el diagnóstico de su estado de conservación. Se caracterizó la estructura diamétrica a partir de 5 cm de amplitud de clases, y la estructura por estado del taxón. Se evaluó la dinámica de la especie a través del registro de todos los individuos por tres años consecutivos (2011-2013), y la determinación de los parámetros tasa media anual de mortalidad, tasa anual de cambios y la tasa finita de crecimiento natural; además se determinó la relación entre abundancia de plántulas y la exposición a la luz mediante un análisis de correspondencia. Se encontró que Juglans jamaicensis presentó una estructura diamétrica irregular que denota reproducción buena y reclutamiento deficiente de sus grupos de árboles en el bosque, sugiriendo cierto desequilibrio entre mortalidad y reclutamiento. El crecimiento y la permanencia de los individuos son los parámetros que tendrían mayor influencia en el mantenimiento de Juglans jamaicensis en el bosque dado que los indicadores tasa media anual de mortalidad, tasa de cambio de la población y tasa finita de crecimiento natural reflejan alta mortalidad en el estadio de plántulas como fase más vulnerable del ciclo de vida de la especie, e indican un reclutamiento y estabilidad poblacional no favorable. Se confirma la tolerancia de las plántulas de Juglans jamaicensis en los dos primeros años de vida.
Palabras clave: Juglans jamaicensis, reclutamiento, estructura, plántula, mortalidad.
ABSTRACT
The work was done in the Turquino National Park in order to analyze the diametric structure, mortality and recruitment of Juglans jamaicensis C. DC., for the diagnosis of its condition. The diametric structure from 5 cm width classes and the taxon state structure was characterized. Dynamics of the species through the registration of all individuals for three consecutive years (2011-2013) was evaluated, and parameters determining the average annual death rate, annual rate of change and the finite rate of natural increase. In addition, the relationship between seedling abundance and exposure to light through a correspondence analysis was determined. It was found that Juglans jamaicensis presents an irregular diametric structure denoting a good reproduction and recruitment failure of its groups of trees in the forest, suggesting some imbalance between mortality and recruitment. The growth and retention of individuals are the parameters that have the greatest influence in maintaining Juglans jamaicensis in the forest since the rate indicators mean annual mortality rate of population change and finite natural growth rate reflect high mortality seedling stage as more vulnerable life cycle of the species phase, indicating a recruitment and population stability is not favorable. Tolerance of seedlings of Juglans jamaicensis in the first two years of life was confirmed.
Key words: Juglans jamaicensis, recruitment, structure, seedling, mortality.
INTRODUCCIÓN
Las especies arbóreas son ecológica, cultural y económicamente valiosos componentes de la biodiversidad y su conservación es fundamental para el bienestar de la población de todos los países del mundo. Con las crecientes presiones de carácter general sobre los ecosistemas y las presiones selectivas sobre las especies, ha resultado evidente que muchas especies arbóreas están amenazadas de extinción. La información sobre el nivel y alcance de la amenaza ha sido sin embargo hasta ahora esparcida y escasa (FAO, 2013).
La mayoría de las especies del género Juglans son exportables para uso maderero. Algunas son puramente forestales, mientras que otras han sido tradicionalmente cultivadas para la obtención de su fruto (Aletá et al., 2003).
Juglans jamaicensis C. DC., es una de las especies más importantes desde el punto de vista maderable y ecológico de las Antillas, por la calidad de su madera y su estado de especie En Peligro Crítico (Berazaín et al., 2005). Esta, cuyo pariente más cercano es Juglans venezuelensis Manning del norte de América del Sur, produce nueces comestibles y una madera muy atractiva similar a la del nogal negro (Juglans nigra L.) de América del Norte, pero su popularidad se ve limitada por su rareza en los bosques húmedos montanos de Cuba, la Española y Puerto Rico (Francis y Alemañy, 1994).
Desde 1923 está prohibida su corta por el decreto Ley 979 del 4 de julio de 1923 (Betancourt, 2000) y la mayor parte de los autores que han referido estudios sobre la especie coinciden en que es gregaria, de escasa aparición en la vegetación y prefiere lugares sombreados en los cañadones, junto a torrentes y barrancos de las regiones montañosas.
Los reportes también refieren la depresión de sus poblaciones, así como la alteración de su hábitat por el cultivo del café en las montañas, su extracción irracional en busca de madera de calidad, a lo cual se le suman los aludes o deslizamientos de suelo y el impacto del cambio climático visto a través del incremento de los eventos meteorológicos extremos como los huracanes (Fors, 1967; Francis y Alemañy, 1994; Risco, 1995; Betancourt, 2000; Bibb y Monsegur, 2013).
En el Parque Nacional Turquino la especie se encuentra formando grupos de pocos árboles con escasa regeneración, y con indicios de extracción antrópica por la calidad de la madera (Rodríguez et al., 2014), así como afectado su vegetación acompañante por el cultivo del café y los deslizamientos de tierra (Lastres et al., 2011).
Por tanto, se define como objetivo analizar la estructura diamétrica, la mortalidad y el reclutamiento de Juglans jamaicensis C. DC., para el diagnóstico de su estado de conservación en el Parque Nacional Turquino.
MATERIAL Y MÉTODO
Ubicación geográfica del área de estudio.
El área de estudio comprendió fragmentos remanentes de bosques de montaña afectados por el cultivo del cafeto y su abandono, así como por los deslizamientos de tierra. Todos los sitios se ubicaron junto a corrientes de agua pequeñas que favorecen alto contenido de humedad en el ambiente. Se georeferenciaron, con un GPS Garmin, siete grupos de árboles de Juglans jamaicensis (Tabla 1) distribuidos en las Unidades Zonales de Conservación Santo Domingo y La Platica.

Metodología empleada
Se caracterizó la estructura horizontal de Juglans jamaicensis mediante un análisis de frecuencias diamétricas con una amplitud de 5 cm. Para caracterizar la estructura por estado de Juglans jamaicensis se utilizó, modificada por el autor, la metodología de Sáenz y Finegan (2000) para estudios de la regeneración natural en bosques tropicales (Tabla 2).
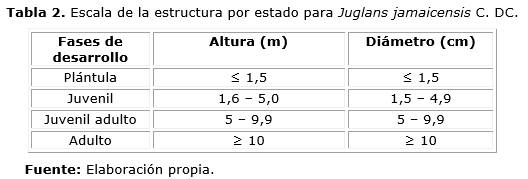
En el caso de las plántulas se realizó un censo, debido a su reducido número en las cercanías de los árboles padres.
La dinámica de Juglans jamaicensis se evaluó a través del registro de todos los individuos de la especie por tres años consecutivos (2011-2013), y calculados los parámetros tasa media anual de mortalidad, tasa anual de cambios y la tasa finita de crecimiento natural.
La tasa media anual de mortalidad se calculó usando la siguiente expresión: TM = (1-((N0-Nm) /N0)1/Δt) *100 (Sheil et al., 1995), donde Nm es el número de individuos muertos de la población inicial, No es el tamaño poblacional en el censo 2011 y Δt es el intervalo de tiempo entre censos.
La tasa anual de cambios en la población o reclutamiento usando el modelo de crecimiento logarítmico estandarizado (Lieberman y Lieberman, 1987 y Condit et al., 1996), fue determinada mediante la siguiente fórmula: r = [ln(N1)-ln(N0)]/Δt, donde: Δ es el intervalo de tiempo entre censos; N0 y N1son los tamaños poblacionales en el censo 2011 y en censo 2013 respectivamente. Además se calculó, a través de la expresión: l(N1/N0)1/Δt, la tasa finita de crecimiento natural-lambda (l) que según Rocha (2000) puede expresar la estabilidad numérica de la población.
Para determinar la relación entre abundancia de plántulas y la exposición a la luz se estableció el criterio de expuesta, medianamente expuesta, y no expuesta; considerando expuesta: aquella plántula con completa exposición a la luz; medianamente expuesta: aquellas con cerca de un 50% de exposición solar, y no expuesta: aquellas sometidas totalmente a la sombra (González, 2005), mediante un análisis de correspondencia realizado con SPSS ver 21.
RESULTADOS Y DISCUSIÓN
La estructura diamétrica de Juglans jamaicensis (Figura 1) fue irregular, indicadora que los individuos plántulas y juveniles se encontraban bajo la sombra de árboles de mayor tamaño y edad, así como que pueden sobrevivir bajo condiciones de menor iluminación como bien refiere Lamprech t (1990) al analizar una estructura diamétrica similar.
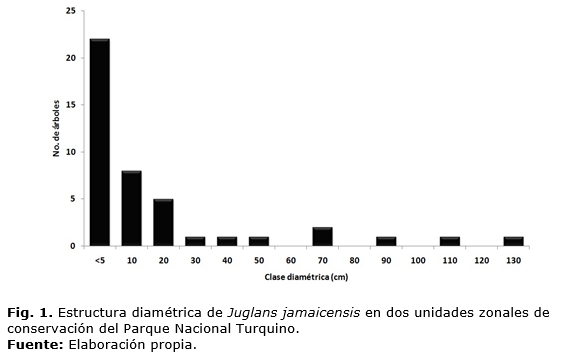
Esta distribución representó la estructura de tipo 3: reproducción buena y reclutamiento malo descrita por Bonger et al., (1988) según Bergmans y Vroomans (1995), aseverando que la mayoría de los individuos se encuentran en las clases de tamaño menores, mientras que las otras clases fueron representadas mínimamente. Asimismo, las irregularidades en la frecuencia de las clases diamétricas sugieren cierto desequilibrio entre la mortalidad y el reclutamiento, también reportados por Pereira y Lima (2010) para varias especies arbóreas del bosque cerrado brasileño de Goiás con igual estructura diamétrica que la encontrada para Juglans en este estudio.
La interpretación de los diámetros en el histograma de frecuencia puede indicar posibles perturbaciones pasadas, naturales o antrópicas como extracción de madera o deforestación ya que Felfili y Silva (1988) exponen que estos eventos aparecen como interrupciones en los histogramas, indicando que el ciclo de vida de la especie no se está completando.
Siendo así las oscilaciones de las clases diamétricas de Juglans jamaicensis sugieren un aumento de la mortalidad de las plantas en el pasado, evidenciado por la baja frecuencia de los individuos de la cuarta a la séptima clase en relación a la clase superior inmediata. Esta irregularidad es indicador de una interrupción en el flujo proporcional de los individuos que ocuparían esas clases intermedias actualmente por el crecimiento de los individuos y apunta hacia la susceptibilidad de la especie a las variaciones y perturbaciones ambientales principalmente de origen antrópico.
Por otro lado, la proporción de individuos en la primera clase indica que tales perturbaciones cesaron y que las poblaciones están en fase de restablecimiento o que como plantean Pereira y Lima (2010) si todavía existen, influencian negativamente a los individuos que se encuentran en el estadio intermediario de desarrollo.
El comportamiento manifestado por esta especie ha sido reportado por Harper (1990) para la mayoría de las especies forestales tropicales de gran porte que presentan individuos bien desarrollados, así como López (2007) para Eschweilera coriacea (DC) S. A. en fragmentos de bosques de la Amazonía brasileña, por Wright et al., (2003) en varias especies en el bosque cerrado de la Isla de Barro Colorado, Panamá, por Keely (1990) para Juglans californica en el sur de California y por Rodríguez et al., (2014) para Juglans jamaicensis en la Jeringa en el Parque Nacional Turquino.
Los resultados obtenidos se corresponden con la distribución discontinua y fragmentada en la Sierra Maestra y en todo el área de distribución natural de la especie en las Antillas (Francis y Alemañy, 1994; Betancourt, 2000), así como el aislamiento y la reducción de sus efectivos poblacionales como consecuencia de las actividades antrópicas incluyendo la sobreexplotación y la agricultura cafetalera en esta zona montañosa (Lastres et al. 2011), lo cual ocasiona, a juzgar por Luna et al., (2006) y Acosta et al, (2011), que en algunas localidades de difícil acceso se observen sólo individuos aislados.
La estructura de Juglans jamaicensis basada en los estadios del ciclo de vida de la especie, mostró inestabilidad en la abundancia de individuos. Las plántulas estuvieron presentes en seis de los siete grupos estudiados, en número reducido denotando aún la capacidad reproductiva de los árboles, sin embargo, los juveniles se concentraron en el 43 % de los grupos (Figura 2), indicando dificultades con el paso de los individuos de plántulas a juveniles.

De igual forma hubo una disminución de los adultos por la pérdida de dos árboles en Palma Mocha producto del impacto del huracán Sandy el 25 de octubre de 2012, durante su azote a la zona oriental del país (Cedeño, 2014). Esto se corresponde con lo expuesto por Lieberman et al., (1985) y Gómes (1998) sobre que la etapa adulta del ciclo de vida de un árbol corresponde a la fase de menor riesgo de muerte para las plantas del bosque húmedo, registrándose de 1 - 4 % de muertes de adultos cada año en las regiones centroamericanas, africanas y asiáticas, causadas por descargas eléctricas, caída de árboles ocasionado por los vientos y cargas excesivas de peso por la presencia de epífitas y lianas.
Varios autores refieren fenómenos similares para especies de este género como Acosta et al., (2011) para Juglans pyriformis Liebm., en México y Francis y Alemañy (1994) y Silander (1999) para Juglans jamaicensis en La Española y Puerto Rico respectivamente donde se reportan poblaciones extremadamente deprimidas (solo 14 individuos en una localidad de la municipalidad de Adjuntas), confirmándose el estado de depresión de las especies de este género en condiciones naturales.
Los principales problemas encontrados en la estructura etárea de Juglans jamaicensis se concentraron en los estadios de plántula y adulto, generando una tasa de mortalidad alta en los mismos, un reclutamiento nulo de los tres primeros estadios y negativo en el estadio de adulto (Tabla 3), lo que favoreció según los valores del, estabilidad en los estadios de plántula, juvenil y juvenil adulto e inestabilidad en el estado de adulto.
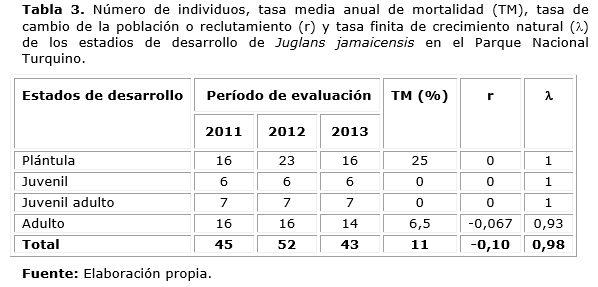
El comportamiento de la tasa de mortalidad evidencia el corto tiempo (un año) en que se reduce la abundancia de plántulas en un 50 %, posiblemente por efectos de la competencia con plantas de mayor crecimiento como han reportado Fredericksen y Mostacedo (2000) para varias especies forestales de bosques bolivianos aprovechados, incluyendo Juglans boliviana.
Martínez (1994) así como Poorter y Hayasido (2000) plantearon al respecto, que la mortalidad de las plántulas es crítica, especialmente cuando hay competencia intra e interespecífica.
El reclutamiento en general (-0,10), fue indicador de la incapacidad de la especie, en el parque, para incrementar su tamaño y la tasa finita de crecimiento natural en el periodo estudiado reafirmó la pérdida de individuos que genera inestabilidad numérica poblacional e inestabilidad en sus estrategias de ocupación de área, lo que asevera que en mayor o menor grado la especie no es viable demográficamente como ha encontrado Rocha (2000) para varias especies en bosque pluvial tropical atlántico de Brasil.
Las diferencias encontradas entre los estadios de desarrollo sugieren la existencia de factores de mortalidad que han afectado la población estudiada. Existen diversos de ellos que podrían explicar la mortalidad de las plántulas.
En la figura 3 se puede apreciar que el número de plántulas en los tres años de evaluación y la exposición a la luz estuvieron asociados por lo que se puede llegar a la conclusión de que la luz es uno de los factores que incide favorablemente en este comportamiento.

En este caso el mayor número de plántulas ocurrió en aquellos lugares con menor exposición a la iluminación en correspondencia con lo planteado por Betancourt (2000) sobre que la especie se regenera naturalmente bastante bien bajo los árboles productores de semilla, así como lo expresado por Francis y Alemañy (1994) y Silander (1999) en cuanto a que cuando las plántulas muestran ese comportamiento, sobreviven por solo un año en el sotobosque sombreado en ausencia de liberación, lo cual se corresponde con este resultado puesto que el número de juveniles es escaso y las plántulas no parecen capaces de sobrevivir por períodos más extensos en posiciones suprimidas.
Al respecto Janzen (1970) planteo que la composición de una comunidad de árboles adultos es en primer lugar una función de la habilidad competitiva de las plántulas y brinzales.
CONCLUSIONES
Juglans jamaicensis presentó en el Parque Nacional Turquino, una estructura diamétrica irregular que denota una reproducción buena y un reclutamiento deficiente de sus grupos de árboles en el bosque, sugiriendo cierto desequilibrio entre mortalidad y reclutamiento.
El crecimiento y la permanencia de los individuos son los parámetros que tendrían mayor influencia en el mantenimiento de Juglans jamaicensis en el bosque dado que los indicadores tasa media anual de mortalidad, tasa de cambio de la población y tasa finita de crecimiento natural reflejan alta mortalidad en el estadio de plántulas como fase más vulnerable del ciclo de vida de la especie, e indican un reclutamiento y estabilidad poblacional no favorable.
Se confirma la tolerancia de las plántulas de Juglans jamaicensis en los dos primeros años de vida.
REFERENCIAS BIBLIOGRÁFICAS
José Luis Rodríguez Sosa. Máster en Ciencias Forestales. Departamento de Ingeniería Forestal, Universidad de Granma. Carretera a Manzanillo Km 17 ½. Peralejo, Bayamo. Granma. Cuba. teléf.: 0123-452249. Correo electrónico: jrodriguezs@udg.co.cu