Revista Cubana de Ciencias Forestales. 2023; enero-abril 11(1): e768. ![]()
Artículo original
Composición, diversidad y estructura arbórea en un bosque de neblina sobre 2 100 msnm en el Perú
Composition, diversity and tree structure in a cloud forest above 2100 masl in Peru
Composição, diversidade e estrutura de árvores em uma floresta nublada acima de 2 100 m de altura no Peru
Josué Otoniel
Dilas-Jiménez1*![]() , Carlos Andrez Mugruza-Vassallo2
, Carlos Andrez Mugruza-Vassallo2![]() , José Luis Marcelo
Peña3
, José Luis Marcelo
Peña3![]()
1Universidad Nacional Autónoma de Tayacaja Daniel Hernández Morillo. Perú.
2Universidad Nacional Tecnológica de Lima Sur. Perú.
3Universidad Nacional de Jaén. Perú.
*Autor para la correspondencia: jdilas@unat.edu.pe
Recibido:2022-08-14.
Aprobado:2023-03-24
RESUMEN
Los bosques de neblina montanos tropicales en el norte del Perú, son severamente fragmentados y degradados por el cambio de uso de la tierra con la consecuente pérdida de importante biodiversidad vegetal. Esta investigación tuvo el objetivo de analizar la composición, diversidad y estructura de la vegetación arbórea en un área de bosque de neblina montano tropical sobre 2 100 m s.n.m en el norte del Perú. En una parcela permanente de una 1 hectárea se marcaron y registraron todos los árboles con diámetros mayores de 10 cm. Se analizó la diversidad, composición florística y estructura de un relicto de bosque neblina. Se registró un total de 792 individuos de 81 especies, 48 géneros y 33 familias. Las familias más ricas en especies fueron Lauraceae (25 especies), Euphorbiaceae (cinco especies), Melastomataceae, Clusiaceae y Rubiaceae (con 4 especies cada una). Cyathea sp1. y Miconia punctata fueron las especies más abundantes y frecuentes. Estos resultados evidencian el alto valor ecológico del bosque estudiado desde el punto de vista de la conservación.
Palabras clave: Montano, neblina, diversidad, florística, estructura.
ABSTRACT
Tropical montane cloud forests in northern Peru are severely fragmented and degraded by land use change with the consequent loss of important plant biodiversity. This research had the objective of analyzing the composition, diversity and structure of the arboreal vegetation in an area of tropical montane cloud forest above 2100 m a.s.l in northern Peru. In a 1-hectare permanent plot, all trees with diametrer over 10 cm were marked and recorded. Diversity, floristic composition and structure of a relict cloud forest were analyzed. A total of 792 individuals of 81 species, 48 genera and 33 families were recorded. The most species-rich families were Lauraceae (25 species), Euphorbiaceae (five species), Melastomataceae, Clusiaceae and Rubiaceae (with 4 species each). Cyathea sp. and Miconia punctata were the most abundant and frequent species. These results show the high ecological value of the forest studied from the point of view of conservation.
Key words: Montane, cloud, diversity, floristic structure.
RESUMO
As florestas tropicais montanas no norte do Peru estão severamente fragmentadas e degradadas pela mudança do uso do solo com a conseqüente perda de importante biodiversidade vegetal. Esta pesquisa teve como objetivo analisar a composição, diversidade e estrutura da vegetação arbórea em uma área de floresta tropical montana acima de 2 100 m a.s.l. no norte do Peru. Em um terreno permanente de 1 hectare, todas as árvores com diâmetro superior a 10 cm foram marcadas e registradas. A diversidade, composição florística e estrutura de uma floresta nublada foram analisadas. Um total de 792 indivíduos de 81 espécies, 48 gêneros e 33 famílias foram registrados. As famílias mais ricas em espécies foram Lauraceae (25 espécies), Euphorbiaceae (cinco espécies), Melastomataceae, Clusiaceae e Rubiaceae (com 4 espécies cada). Cyathea sp1. e Miconia punctata foram as espécies mais abundantes e freqüentes. Estes resultados mostram o alto valor ecológico da floresta estudada do ponto de vista da conservação.
Palavras-chave: Montana, floresta nublada, diversidade, florística, estrutura.
INTRODUCCIÓN
Los bosques de neblina montanos tropicales son ecosistemas de alta importancia en el mundo, abarcan alrededor de 380 000 km2 que representa un 2,5 % de los conocidos bosques tropicales (Rubb et al., 2004), esta importancia viene dada por su excepcional diversidad de flora que albergan al demostrar una alta riqueza de especies por unidad de área, siendo fuente importante de endemismos.
Estos se comportan como eficientes ecosistemas en cuanto a servicios ecosistémicos como la captura de carbono (Álvarez-Arteaga et al., 2013; Eller et al., 2020; Knowles et al., 2020) tanto en su biomasa arbórea y principalmente en el suelo, siendo superado sólo por ecosistemas altoandinos (páramos-puna) con alta concentración de carbono orgánico (Dilas-Jiménez y Huamán Jiménez, 2020; Zimmermann et al., 2010). Sin embargo, estos bosques se ubican en las zonas más sensibles con anomalías climáticas a diferencia de otros ecosistemas terrestres en el mundo (Seddon et al., 2016) y son amenazados con los efectos del cambio climático y principalmente son fragmentados y degradados con el cambio de uso de la tierra (Moreira et al., 2021).
En el Perú, los bosques de neblina montanos tropicales se ubican mayormente desde 1 000 hasta 3 000 m s.n.m (Van de Weg et al., 2014), en la zona norte peruana principalmente se ubican en las regiones de Cajamarca, Piura y amazonas, regiones con amplia diversidad florística al alrededor de 17 000 especies, 800 de estas endémicas (Sagástegui et al., 2003).
En la región Cajamarca, los bosques de neblina montanos tropicales se ubican principalmente en los territorios de las provincias de Jaén y San Ignacio (MINAM, 2014), los cuales por albergar especies maderables con alta demanda como Retrophyllum rospigliosii, Prumnopitys harmsiana (podocarpáceas) están sufriendo tala selectiva, además están amenazados por las altas tasas de deforestación en la zona alrededor 4 000 hectáreas año-1 (Llerena et al., 2010). No obstante, hay escasa información sobre aspectos ecológicos e hidroecológicos que brinda la flora arbórea en estos tipos de bosque, en general, son muy escasos los estudios realizados y publicados para este tipo de bosques en el norte del Perú (Sagástegui et al., 2003; Seddon et al., 2016).
En la actualidad, Perú impulsa estudios sobre diversidad del componente arbóreo principalmente en el bosque amazónico, basados en parcelas de muestreo con dimensión mínima de una hectárea (de área contigua) y metodologías estandarizadas (Marcelo-Peña y Reynel, 2014). Sin embargo, para los bosques de neblina en la zona norte son muy pocos los registros de estudios publicados, entre ellos uno realizado por Peña y Pariente (2015) en un bosque de neblina en la provincia de San Ignacio, Cajamarca, a una altitud de 2 150 m s.n.m registró 308 individuos distribuidos en 31 familias, 30 géneros y 39 especies, siendo las cinco familias más con mayor índice de valor de importancia, en orden descendente, Podocarpaceae, Lauraceae, Rubiaceae, Melastomataceae y Clusiaceae. Marcelo-Peña y Arroyo (2013) en estudio realizado en bosque de neblina de la provincia de Jaén, Cajamarca, sobre los 2 100 msnm registró dos nuevas especies para la ciencia de la familia Magnoliaceae, Magnolia jaenensis cuyas primeras colectas se hicieron con este estudio y M. manguillo.
Ante ello, el presente estudio tuvo el objetivo de analizar la composición, diversidad y estructura de la vegetación arbórea en un área de bosque de neblina montano tropical sobre 2 100 m s.n.m en el norte del Perú.
MATERIALES Y MÉTODOS
Área de estudio
La investigación se desarrolló en una zona de relictos de bosque de neblina montano tropical, a una altitud promedio de 2 170 m s.n.m, localizado en la región Cajamarca, provincia de Jaén, distrito de Huabal, en el caserío San Luis del Nuevo Retiro, ubicado en las coordenadas UTM 0728564 este y 9372902 norte, zona 17M (Datum WGS 84). El sitio se ubica en el área de amortiguamiento del Área de Conservación Municipal Bosques de Huamantanga (ACM-BH).
El área de estudio, siguiendo el criterio de Zonas de Vida (Holdridge, 1978), pertenece a un bosque muy húmedo-Montano Bajo Tropical (bmh-MBT), con pendientes pronunciadas y de fisiografía ondulada, las temperaturas van desde los 12º C hasta los 17º C, y precipitaciones anuales entre 1900 mm y 3800 mm. Los suelos se caracterizan por ser ácidos y abundante materia orgánica (OIKOSlab, 2013 citado por Peña y Pariente, 2015) (Figura 1).
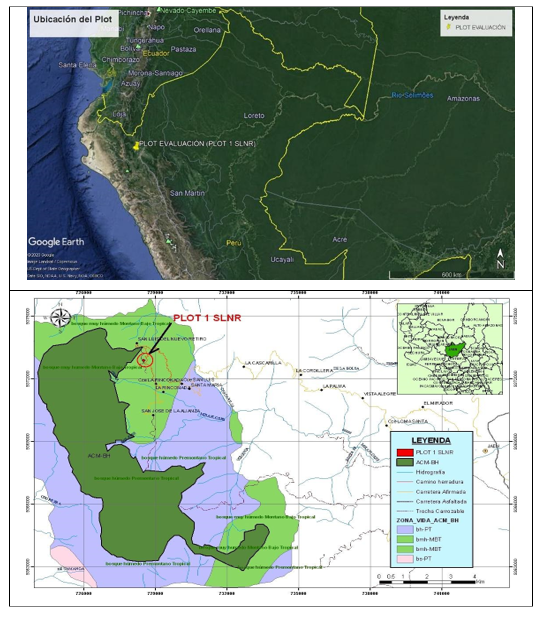
Figura 1. - Superior: ubicación de la parcela permanente en el norte del Perú. Inferior: Ubicación de la parcela en la provincia de Jaén, Cajamarca
Instalación de la parcela de evaluación
Para la instalación de la parcela permanente se siguió la metodología usada por diversos autores RAINFOR, 2016; Synnott, 1979). La selección del sitio para la instalación fue un relicto de bosque de propiedad particular a fin de salvaguardar la conservación de la parcela para futuras remediciones, así como por su estado de conservación en cuanto a la flora existente. La localización de la parcela, el plaqueado y registro de árboles mayores o iguales a 10 cm de diámetro a la altura del pecho (DAP), así como de las primeras colecciones botánicas se realizó en el primer semestre del 2008 (Figura 2).
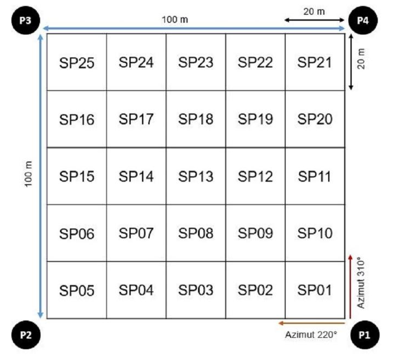
Figura 2. - Instalación y distribución de las subparcelas dentro de la parcela permanente
Colección botánica y registro de información
La colección botánica se desarrolló en dos momentos, la primera colección se ejecutó en el primer semestre 2008 y la segunda colección, al segundo semestre del mismo año, esto principalmente para las muestras que fueron colectadas como estériles en la primera colección, a fin de facilitar el trabajo de identificación; este trabajo se realizó utilizando el equipo estándar, así como los materiales y procedimientos recomendados para este tipo de trabajo (Rodríguez y Rojas, 2006). Para la herborización de las muestras colectadas se siguió metodología estandarizada en estos casos (Rodríguez y Rojas, 2006; Rotta et al., 2008). Los especímenes colectados fueron prensados, previa preservación con una solución de 60% de agua y 40 % de alcohol de 96º y trasladados desde campo hasta la ciudad de Jaén donde se hicieron los procedimientos de secado y montaje de muestras (Marcelo-Peña et al., 2011; Rotta et al., 2008). Los especímenes colectados, secados y montados fueron enviados al Herbario MOL de la Universidad Nacional Agraria La Molina para su identificación taxonómica.
Manejo y análisis de información
Para el manejo ordenado de datos, en campo se codificó con placas de aluminio a cada individuo censado dentro de la parcela permanente, para lo cual se utilizó códigos numéricos de 6 dígitos, siendo este mismo el código de colección, por ejemplo, código 108220, haciendo la lectura de izquierda a derecha, el primer número indica el número de Parcela (Parcela 1), los siguientes dos números indican el número de subparcela dentro de la parcela permanente (subparcela 08) y finalmente los tres últimos números indican el número del individuo (individuo 220).
Con el fin de conocer la efectividad del muestreo, en la medición de individuos realizados por subparcela se llevó el conteo del número de especies que incrementaron a medida que se incrementó el número de subparcelas analizadas, con lo cual se elaboró un gráfico de curva área-especie (Matteucci y Colma, 1982; Chu, Smith y Solow, 2014).
Con la información de la identificación taxonómica recibida del Herbario MOL, se procedió al análisis de las siguientes variables (Antón y Reynel, 2004; Matteucci y Colma, 1982):
Diversidad: se determinó el número de individuos por ha, número de familias, número de especies. Asimismo, se determinó los siguientes índices de diversidad (Ecuación 1); (Ecuación 2); (Ecuación 3) y (Ecuación 4):
Riqueza específica (S):
![]()
Índice de dominancia de Simpson (D):
![]()
Índice de diversidad de Simpson (1-D, Simpson, 1949):
![]()
Índice de equidad de Shannon-Wiener (H, Hill, 1973):
![]()
Composición: se determinó las familias y géneros más abundantes, especies endémicas y raras.
Variables dasométricas: se determinó el área basal (m2), altura total (m).
Estructura de la vegetación: se calculó la Densidad o Abundancia, la Frecuencia y la Dominancia de las especies identificadas. Con estos tres datos se obtuvo el Índice de Valor de Importancia (IVI) del total de especies, este índice permite medir cómo las especies contribuyen en la estructura de un ecosistema (Cottam y Curtis, 1956; Ragavan et al., 2015).
RESULTADOS Y DISCUSIÓN
Composición florística
Se registró 792 individuos para la parcela permanente de 1ha, pertenecientes a 81 especies de 48 géneros y 33 familias. El cociente de mezcla en este estudio es de 0,10 (81/792) es decir existen unos 10 individuos por cada especie, este resultado es similar a los encontrados en estudios en selva central del Perú (Marcelo-Peña y Reynel, 2014), pero inferior a otros estudios realizados en años anteriores en estas mismas zonas donde los cocientes de mezcla estaban alrededor de 0,22 (Antón y Reynel, 2004). El bajo coeficiente de mezcla encontrado estaría vinculado a la alta proporción de individuos de dos especies Cyathea sp. y Miconia punctata que representaron el 48 % del total.
En al análisis de la curva área-especie, del total de especies registradas (81), se encontró que el 67 % de estas se alcanzó a completar en la subparcela 9 y el 85 % al completar la subparcela 19 (Figura 3).
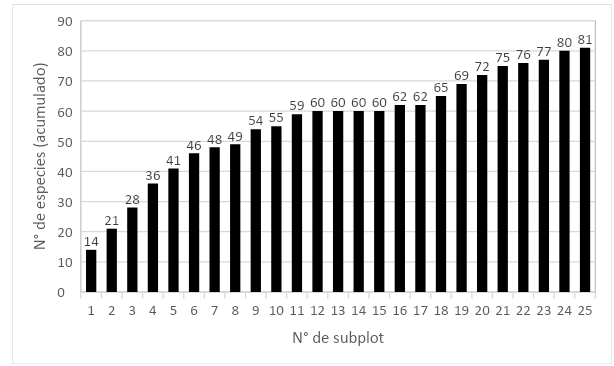
Figura 3. - Curva área-especie en la parcela permanente evaluada
Diversidad florística
La familia Lauraceae (25 especies) es la más rica en especies, seguida de Euphorbiaceae (cinco especies), Melastomataceae, Clusiaceae y Rubiaceae (con cuatro especies cada una). La familia Lauraceae es endémica en estos bosques montanos en el Perú, con mayor abundancia en sus géneros Ocotea y Nectandra (León, 2006), así también presencia importante de la familia Melastomataceae en su género Miconia (Ledo et al., 2012).
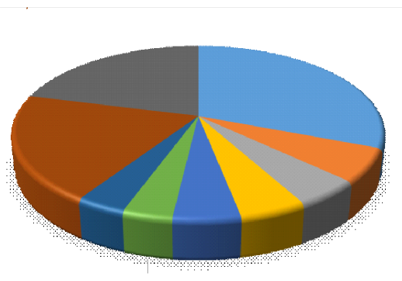
Figura 4. - Abundancia relativa de especies por familia en el bosque montano de neblina, Jaén, Cajamarca
En evaluación realizada en los bosques de Chinchiquilla, San Ignacio, bosque montano de neblina a 2 150 m s.n.m, se encontró similares resultados en cuanto a las cinco familias con mayor número de individuos como Melastomataceae y Rubiaceae, y al cociente de mezcla encontrado en esta investigación, sin embargo, se evidencia una diferencia saltante debido a que en estos bosques de San Ignacio resalta en número las podocarpáceas o árboles conocidos en la zona como "romerillos" (Peña y Pariente, 2015).
En cuanto a la diversidad alfa según los índices analizados, donde una población sería homogénea si la distribución espacial entre especies es uniforme en la zona de estudio (Matteucci y Colma, 1982), los resultados para el presente estudio son: Riqueza específica (S), con un valor de 81 especies; Índice de dominancia de Simpson (D), con un valor de 0,138; Índice de diversidad de Simpson (1-D), con un valor de 0,862; y el Índice de equidad de Shannon-Wiener (H), con un valor de 2,856. Así, considerando al Índice de Shannon como un indicador de homogeneidad de un ecosistema evaluado (Shi y Zhu, 2009), el bosque evaluado en el presente estudio mostró valores intermedios, similares a los encontrados en un parcela permanente instalada en la misma zona sobre 2500 msnm, (Pérez, 2011), así también similares a resultados de evaluación en el Fundo Génova, Chanchamayo, en tipo de bosque secundario tardío (Antón y Reynel, 2004), pero inferiores a los valores de este índice (3,309) encontrados para el bosque un bosque montano de neblina en Chinchiquilla, San Ignacio, Cajamarca (Peña y Pariente, 2015) e inferiores a resultados de otros bosques premontanos y montanos en La Merced, San Ramón y Satipo en Perú (Marcelo-Peña y Reynel, 2014).
Endemismo
Al confrontar la lista de especies con el Catálogo de las Angiospermas y Gimnospermas del Perú (Brako y Zarucchi, 1993); se registró seis especies endémicas estas son:
Cabe precisar que se encontró individuos de Podocarpus oleifolius (Podocarpaceae), que es la única familia de coníferas nativas del Perú y que es propia de los bosques nublados en las provincias de Jaén y San Ignacio en Cajamarca, en parcela permanente instalada en los bosques de Chinchiquilla en San Ignacio se encontró que estas podocarpáceas (Prumnopitys harmsiana y Podocarpus glomeratus) representan un 12% del total de individuos (Peña y Pariente, 2015). Estas podocarpáceas son protegidas en el Santuario Nacional Tabaconas Namballe (SNTN) pero son altamente amenazadas por la tala indiscriminada por ser una especie maderable muy requerida en la zona (Elliot, 2009; Llerena et al., 2010).
Asimismo, se destaca la presencia de un individuo que en este estudio se identificó preliminarmente como especie del género Talauma (Magnoliaceae), conocido como "militar. Esto fue base para estudios posteriores en la zona, principalmente en alrededores del poblado de San Luis del Nuevo Retiro, donde efectivamente se confirmó la existencia de especies de Magnoliaceas nuevas para la ciencia, reportándose así dos especies nuevas nombradas como Magnolia jaenensis y M. manguillo, estas nuevas especies a la vez corresponden a primeros registros de Magnolia en bosques montanos sobre los 2 100 m s.n.m (Marcelo-Peña y Arroyo, 2013).
No se descarta que pudiera haber otras especies endémicas o nuevas para la flora peruana e incluso para la ciencia; debido a que varios de los especímenes colectados, fueron registrados sin órganos reproductivos (flores y/o frutos), dificultando la identificación taxonómica de los especímenes hasta el nivel de especie.
Estructura de la vegetación (abundancia, frecuencia y dominancia)
De los 792 individuos censados, las cinco especies más abundantes y que representan un 63% del total de individuos son Cyathea sp. (30,93 %), Miconia punctata (17,17 %), Helicostylis tovarensis (6,69 %), Myrcia sp. (4,80 %) y Hedyosmum angustifolium (3,66 %), ver detalle en el apéndice 1.
Los cinco géneros más abundantes en orden descendente son Cyathea (Pteridophyta) con 245 individuos (30,93 %), Miconia (Melastomataceae) con 140 individuos (17,68 %), Helicostylis (Moraceae) con 53 individuos (6,69 %), Myrcia (Myrtaceae) con 42 individuos (5,30 %) y Ocotea (Lauraceae) con 32 individuos (4,04 %).
Así también las cinco más abundantes en orden son Cyatheaceae (31,31 %), Melastomataceae (18,31 %), Lauraceae (7,58 %), Moraceae (7,20 %) y Myrtaceae (5,30 %). Cyatheaceae fue la familia más abundante en el sitio estudiado. Estas familias son propias de estos tipos de bosques montanos en América (Ledo et al., 2012; Schin-ichiro y Kitayama, 1999), incluso algunas de estas también presentes en otros continentes (Shi y Zhu, 2009).
En cuanto a la frecuencia y dominancia, estas variables guardan estrecha relación con la abundancia de las especies. En la Tabla 1, se muestra el Índice de Valor de Importancia (IVI) para las diez especies más importantes (ver detalle en apéndice 1), donde se puede ver que sólo dos especies Cyathea sp. y Miconia punctata representan cerca de la tercera parte del IVI total.
Tabla 1. - Índice de Valor de Importancia de las diez especies más importantes del bosque estudiado
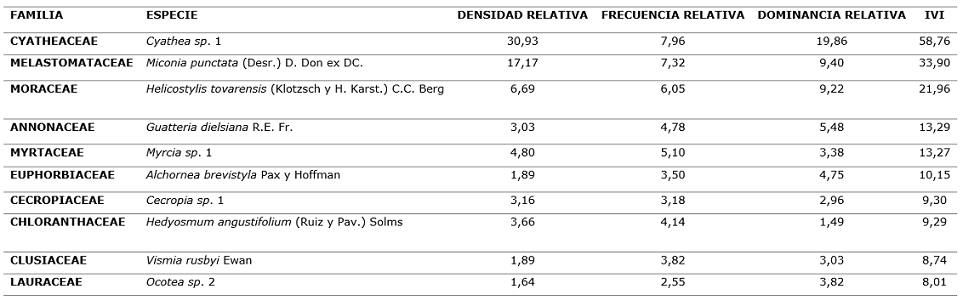
La notable abundancia de la Cyatheaceae en el bosque estudiado es debida a que esta familia es representativa de los bosques montanos, se reconocen al menos unas 83 especies del género Cyathea en el Perú (Lehnert, 2011).
Estos resultados también muestran similitud con resultados de otros estudios en ecosistemas similares en Selva Central del Perú, donde también se encontró la dominancia del género Miconia de las melastomatáceas (Lehnert, 2011), aunque en este estudio si bien es una de las familias más abundantes, es mucho menos diversa que las lauráceas (Figura 4).
De acuerdo al análisis de Densidad, Frecuencia y Dominancia de todas especies encontradas en la parcela permanente, el Índice de Valor de Importancia (IVI) encontrado para las cinco primeras especies, en orden decreciente es: Cyathea sp 1 (58,76), Miconia punctata (33,90), Helicostylis tovarensis (21,96), Guatteria dielsiana (13,29) y Myrcia sp 1 (13,27).
Estos resultados difieren en cuanto a su nivel de importancia para las cinco primeras especies encontradas en los bosques montanos de neblina en la zona de San Ignacio, Cajamarca, donde las especies Prumnopitys harmsiana y Podocarpus glomeratus (podocarpáceas) son las más importantes, seguidas de Cinchona sp., Cecropia sp. y Endlicheria sp. (Peña y Pariente, 2015). Especies del género Miconia (melastomataceae), especies del género Guatteria (annonaceae) y especies de Moraceas se reportan entre las primeras 10 especies de importancia en bosques montanos y premontanos en el Perú (Antón y Reynel, 2004; Marcelo-Peña y Reynel, 2014; Peña y Pariente, 2015). En el Apéndice 1, se muestra el Índice de Valor de Importancia (IVI) del total de las especies.
Análisis las variables dasométricas y volumétricas las especies
De todos los individuos censados dentro, el diámetro (DAP) promedio fue 19,64 cm, con una varianza de 90,5; el área basal total es 29,62 m2, el área basal promedio por individuo es 0,037 m2, con una varianza de 0,002; la altura total de los árboles tiene un promedio de 10,53m, con una varianza de 13,86; y la altura comercial tiene un promedio de 6,72 m, con una varianza de 5,19. En evaluación realizada en la misma zona boscosa pero a una altitud de 2543 msnm se encontró un DAP promedio de 16,44 cm (Pérez, 2011) lo cual indica que a mayor altitud los diámetros de los árboles reducen, mientras que en otro bosque de neblina en San Ignacio, Cajamarca cuya Parcela permanente se instaló a una altura similar al presente estudio se encontró un DAP promedio de 25.20 cm, este mayor diámetro fue influenciado por la alta presencia de individuos de la familia Podocarpaceae que presentaron las mayores altura y diámetros, pues corresponden a especies forestales maderables de la zona (Peña y Pariente, 2015).
CONCLUSIONES
Los bosques de neblina montanos tropicales ubicados en la zona alta de la provincia de Jaén tienen importante diversidad de especies, así como especies endémicas y potencialidad de albergar especies nuevas para la ciencia.
La familia Lauraceae con 15 especies registradas fue la familia más diversa en el bosque estudiado, confirmándose así que las lauráceas son una de las familias más diversas en los bosques montanos en el Perú.
En cuanto a la estructura vegetal del bosque estudiado, la alta abundancia de la familia Cyatheaceae y, así como de la1familia Melastomataceae confirma la representatividad de especies de estas familias en los bosques montanos de neblina.
REFERENCIAS BIBLIOGRÁFICAS
ÁLVAREZ-ARTEAGA, G., CALDERÓN, N. E. G., KRASILNIKOV, P., Y GARCÍA-OLIVA, F. 2013. Almacenes de carbono en bosques montanos de niebla de la Sierra Norte de Oaxaca, México. Agrociencia, vol. 47 no. 2, pp. 171-180. Disponible en: https://www.scielo.org.mx/scielo.php?pid=S1405-31952013000200006yscript=sci_abstractytlng=pt
ANTÓN, D., Y REYNEL, C. 2004. Relictos de bosques de excepcional diversidad en los Andes Centrales del Perú (UNALM (ed.); Primera Ed). APRODES/Asociación Peruana para la Promoción del Desarrollo Sostenible. Disponible en: http://infobosques.com/descargas/biblioteca/446.pdf
BRAKO, L., Y ZARUCCHI, J. L. 1993. Catalogue of the flowering plants and gymnosperms of Peru: Catálogo de las angiospermas y gimnospermas del Perú. Monographs in Systematic Botany from the Missouri Botanical Garden, 45, 11286.
CHU, C., SMITH, W. y SOLOW, A. 2014. A hidden species-area curve. Environ Ecol Stat 21, pp. 113-124. DOI. https://doi.org/10.1007/s10651-013-0247-2
COTTAM, G., Y CURTIS, J. T. 1956. The Use of Distance Measures in Phytosociological Sampling. Ecology, vol. 37 no. 3, pp. 451-460. DOI. https://doi.org/https://doi.org/10.2307/1930167
DILAS-JIMÉNEZ, J. O., Y HUAMÁN JIMÉNEZ, A. O. 2020. Captura de carbono por un bosque montano de neblina del Perú. Revista de Investigación Científica y Tecnológica Alpha Centauri, vol. 1 no. 3, pp. 13-25. DOI. https://doi.org/https://doi.org/10.47422/ac.v1i3.16
ELLER, C. B., MEIRELES, L. D., SITCH, S., BURGESS, S. S. O., Y OLIVEIRA, R. S. 2020. How Climate Shapes the Functioning of Tropical Montane Cloud Forests. Current Forestry Reports, vol. 6 no. 2, pp. 97-114. DOI. https://doi.org/10.1007/s40725-020-00115-6
ELLIOT, J. 2009. Los bosques de la cuenca transfronteriza del rio Mayo-Chinchipe Perú-Ecuador (ITDG (ed.); Primera Ed). Soluciones Prácticas ITDG. Disponible en: http://infobosques.com/portal/wp-content/uploads/2017/09/Los-bosques-de-la-cuenca-transfronteriza-del-río-Mayo-Chinchipe.pdf
Hil, H.l 1973. Diversity and Evenness: A Unifying Notation and Its Consequences. Ecology, vol. 54 no. 2, pp. 427-432. Doi:10.2307/1934352
HOLDRIDGE, L. R. 1978. Ecología basada en zonas de vida. IICA, Instituto Interamericano de Ciencias Agrícolas 1978 - 216 pp. Disponible en:.https://books.google.com.cu/books?id=Sg1oK3Zhn10C&printsec=copyright#v=onepage&q&f=false
KNOWLES, J. F., SCOTT, R. L., BIEDERMAN, J. A., BLANKEN, P. D., BURNS, S. P., DORE, S., KOLB, T. E., LITVAK, M. E., Y BARRON-GAFFORD, G. A. 2020. Montane forest productivity across a semiarid climatic gradient. Global Change Biology, August, pp. 114. DOI. https://doi.org/10.1111/gcb.15335
LEDO, A., CONDÉS, S., Y ALBERDI, I. 2012. Forest biodiversity assessment in Peruvian Andean Montane cloud forest. Journal of Mountain Science, vol. 9 no. 3, pp. 372-384. DOI. https://doi.org/10.1007/s11629-009-2172-2
LEHNERT, M. 2011. The Cyatheaceae (Polypodiopsida) of Peru. Brittonia, vol. 63 no. 1, pp. 11-45. DOI. https://doi.org/10.1007/s12228-009-9112-x
LEÓN, B. 2006. Lauraceae endémicas del Perú. Revista Peruana de Biología, vol. 13, no. 2, pp. 669-677. Disponible en: http://www.scielo.org.pe/scielo.php?script=sci_arttextypid=S1727-99332006000200065
LLERENA, C., CRUZ-BURGA, Z., DURT, É., MARCELO-PEÑA, J., MARTINEZ, K., Y OCAÑA, J. 2010. Gestión ambiental de un ecosistema frágil. Los bosques nublados de San Ignacio, Cajamarca, cuenca del rio Chinchipe (ITDG (ed.); Primera Ed). Soluciones Prácticas ITDG. Disponible en: http://infobosques.com/portal/wp-content/uploads/2017/09/Gestión-ambiental-de-un-ecosistema-frágil.pdf
MARCELO-PEÑA, J.L., Y ARROYO, F. 2013. Magnolia jaenensis y M. manguillo, nuevas especies de Magnoliaceae del norte de Perú. Brittonia, vol. 65, no. 1, pp. 106-112. DOI. https://doi.org/10.1007/s12228-012-9280-y
MARCELO-PEÑA, J.L., Y REYNEL, C. 2014. Patrones de diversidad y composición florística de parcelas de evaluación permanente en la selva central de Perú. Rodriguesia, vol. 65, no. 1, pp. 35-47. Disponible en: http://rodriguesia-seer.jbrj.gov.br/index.php/rodriguesia/article/view/735
MARCELO-PEÑA, JOSÉ L., REYNEL RODRÍGUEZ, C., Y ZEVALLOS, P. (2011). Manual de Dendrología. CONCYTECEditor: JL Marcelo Peña ISBN: 978-9972-50-131-9. Disponible en: https://www.researchgate.net/publication/266560129_Manual_de_Dendrologia
MATTEUCCI, S., Y COLMA, A. 1982. Metodología para el estudio de la vegetación. Secretaría General de la Organización de los Estados Americanos. Disponible en: https://aulavirtual.agro.unlp.edu.ar/pluginfile.php/76505/mod_resource/content/3/MatteucciColma1982.pdf
MINAM. 2014. Perú reino de bosques (P. Editores (ed.); Primera Ed). Ministerio del Ambiente. Disponible en: http://www.bosques.gob.pe/archivo/1455ad_perureinodebosques.pdf
MOREIRA, B., VILLA, P. M., ALVEZ-VALLES, C. M., Y CARVALHO, F. A. 2021. Species composition and diversity of woody communities along an elevational gradient in tropical Dwarf Cloud Forest. Journal of Mountain Science, vol 18, no. 6, pp. 1489-1503. DOI. https://doi.org/10.1007/s11629-020-6055-x
PEÑA, G., Y PARIENTE, E. 2015. Composición y diversidad arbórea en un área de bosque de Chinchiquilla, San Ignacio-Cajamarca, Perú. Arnaldoa, vol 22, no 1, pp. 139-154. Disponible en: http://200.62.226.189/Arnaldoa/article/viewFile/187/175
RAGAVAN, P., SAXENA, A., MOHAN, P. M., RAVICHANDRAN, K., JAYARAJ, R. S. C., Y SARAVANAN, S. 2015. Diversity, distribution and vegetative structure of mangroves of the Andaman and Nicobar Islands, India. Journal of Coastal Conservation, vol 19, no. 4, pp. 417-443. DOI. https://doi.org/10.1007/s11852-015-0398-4
RAINFOR. 2016. Manual de Campo para el Establecimiento y Remedición de Parcelas. Disponible en: http://www.rainfor.org/upload/ManualsSpanish/Manual/RAINFOR_field_manual_version2016_ES.pdf
RODRÍGUEZ R., E. F., Y ROJAS G., R. P. 2006. El herbario. Administración y manejo de colecciones botánicas. ISSUU. Disponible en: https://issuu.com/ericrodriguezr/docs/herbario
ROTTA, E., DE CARVALHO, L. C., Y ZONTA, M. 2008. Manual de Prática de Coleta e Herborização de Material Botânico (EMBRAPA (ed.); 1a edição). Dados Internacionais de Catalogação na Publicação (CIP) Embrapa Florestas. Disponible en: https://www.infoteca.cnptia.embrapa.br/infoteca/handle/doc/315636
RUBB, P., MAY, L., MILES, L., Y SAYER, J. 2004. Cloud forest agenda (UNEP (ed.); First Edit). United Nations Environment Programme-UNEP. Disponible en: https://www.researchgate.net/publication/318657347_Cloud_Forest_Agenda
SAGÁSTEGUI, A., SÁNCHEZ, I., ZAPATA, M., Y DILLON, M. 2003. Diversidad florística del norte del Perú, Tomo II bosques montanos (GRAFICART (ed.). GRAFICART. Disponible en: https://www.nhbs.com/diversidad-floristica-del-norte-de-peru-volume-2-book
SCHIN-ICHIRO, A., Y KITAYAMA, K. 1999. Structure, Composition and Species Diversity in an Altitude-Substrate Matrix of Rain Forest Tree Communities on Mount Kinabalu, Borneo Author (s): Shin-ichiro Aiba and Kanehiro Kitayama Structure, mat. Plant Ecology, vol, 140, no 2, pp. 139-157. Stable URL: http://www.jstor.org/stable/20050735 DOI. https://doi.org/https://doi.org/10.1023/A:1009710618040
SEDDON, A. W. R., MACIAS-FAURIA, M., LONG, P. R., BENZ, D., Y WILLIS, K. J. 2016. Sensitivity of global terrestrial ecosystems to climate variability. Nature, 531, pp. 65-72. Disponible en: https://doi.org/https://doi.org/10.1038/nature16986
SHI, J. P., Y ZHU, H. 2009. Tree species composition and diversity of tropical mountain cloud forest in the Yunnan, southwestern China. Ecological Research, vol 24, no 1, pp. 83-92. DOI. https://doi.org/10.1007/s11284-008-0484-2
SIMPSON, E. .1949. Measurement of diversity. Nature 163688. Disponible en: https://doi.org/10.1038/163688a0
SYNNOTT, T. J. 1979. Manual of permanent plot procedures for tropical rainforests. Tropical Forestry Papers. Oxford University. Disponible en: https://ora.ox.ac.uk/objects/uuid:c1ab9dff-f60a-4d9b-9b3d-5706264b1d4d/download_file?file_format=pdfysafe_filename=TFP14.pdfytype_of_work=Working+paper
VAN DE WEG, M. J., MEIR, P., WILLIAMS, M., GIRARDIN, C., MALHI, Y., SILVA-ESPEJO, J., Y GRACE, J. 2014. Gross Primary Productivity of a High Elevation Tropical Montane Cloud Forest. Ecosystems, vol, 17, no 5, pp. 751-764. DOI. https://doi.org/10.1007/s10021-014-9758-4
ZIMMERMANN, M., MEIR, P., SILMAN, M. R., FEDDERS, A., GIBBON, A., MALHI, Y., URREGO, D. H., BUSH, M. B., FEELEY, K. J., GARCIA, K. C., DARGIE, G. C., FARFAN, W. R., GOETZ, B. P., JOHNSON, W. T., KLINE, K. M., MODI, A. T., RURAU, N. M. Q., STAUDT, B. T., Y ZAMORA, F. 2010. No differences in soil carbon stocks across the tree line in the Peruvian Andes. Ecosystems, vol 13 no. 1, pp. 62-74. DOI. https://doi.org/10.1007/s10021-009-9300-2
Conflicto de intereses:
Los autores declaran no tener conflictos de intereses.
Contribución de los autores:
Los autores han participado en la redacción del trabajo y análisis de los documentos.
![]()
Esta obra está bajo una licencia de Creative Commons Reconocimiento-NoComercial
4.0 Internacional.
Copyright (c) 2023
Josué Otoniel
Dilas-Jiménez, Carlos Andrez Mugruza-Vassallo, José Luis Marcelo
Peña