Revista Cubana de Ciencias Forestales. 2021; septiembre-diciembre 9(3): 395-411 ![]()
Efecto del sustrato y la micorriza arbuscular en el sistema radical y estado nutricional de Swietenia mahagoni L. Jacq.
Effect of substrate and arbuscular mycorrhiza on the root system and nutritional status of Swietenia mahagoni L. Jacq.
Efeito do substrato e da micorriza arbuscular no sistema radicular e estado nutricional da Swietenia mahagoni L. Jacq.
Emir Falcón Oconor1*![]() https://orcid.org/0000-0001-8833-4942
https://orcid.org/0000-0001-8833-4942
Milagros Cobas López2![]() https://orcid.org/0000-0002-3785-5235
https://orcid.org/0000-0002-3785-5235
Marta Bonilla Vichot2![]() https://orcid.org/0000-0002-6605-5296
https://orcid.org/0000-0002-6605-5296
Orfelina Rodríguez Leyva1![]() https://orcid.org/0000-0002-1575-1515
https://orcid.org/0000-0002-1575-1515
1Universidad de Guantánamo.
Guantánamo, Cuba.
![]()
2Universidad de Pinar del Río "Hermanos Saíz Montes de Oca". Pinar del Río, Cuba.![]()
*Autor para la correspondencia: emir@cug.co.cu
Recibido: 26/04/2020.
Aprobado: 19/10/2021.
RESUMEN
El estudio se desarrolló con el objetivo de evaluar la influencia de los factores sustratos y cepas de hongos micorrízicos arbusculares (HMA) sobre el sistema radical y estado nutricional de Swietenia mahagoni L. Jacq. Los factores probados fueron, factor 1 sustrato: cascarilla de cacao, fibra de coco y aserrín de pino compostado en proporciones 6:2:2 y 2:6:2, y un testigo compuesto por suelo al 100 %; factor dos cepas micorrízicas: Glomus cubense, Rhizoglomus irregulare y Funneliformis mosseae. Los nueve tratamientos resultantes del arreglo factorial 3x3, se establecieron bajo un diseño experimental completamente al azar. Fueron medidas las variables relacionadas con el sistema radical (largo de la raíz principal, cantidad de raíces primarias y secundarias, colonización micorrízica) y acumulación de nitrógeno, fósforo y potasio mediante un análisis gráfico de vectores. Los resultados obtenidos demostraron que existió un efecto diferenciado de las micorrizas en el crecimiento de S. mahagoni. La inoculación con las cepas micorrízicas mejoró la nutrición de las plantas. El mejor sustrato para el establecimiento de la asociación micorrízica fue el S2 conformado por 20 % de cascarilla de caco + 60 % de fibra de coco + 20 % de aserrín de pino, pero los mejores tratamientos para el crecimiento y nutrición de las plántulas fueron la combinación del sustrato S2 y las cepas Glomus cubense y Rhizophagus irregulare.
Palabras clave: Análisis de vectores; Colonización micorrízica; Nutrición forestal; Sistema radical.
ABSTRACT
The study was developed with the objective of evaluating the influence of substrate factors and strains of arbuscular mycorrhizal fungi (AMF) on the root system and nutritional status of Swietenia mahagoni L. Jacq. The factors tested were: factor 1 substrate: cocoa husk, coconut fiber and composted pine sawdust in proportions 6:2:2 and 2:6:2 and a control composed of 100 % soil; factor 2 mycorrhizal strains: Glomus cubense, Rhizoglomus irregulare and Funneliformis mosseae. The nine treatments resulting from the 3x3 factorial arrangements were established under a completely randomized experimental design. Variables related to the root system (length of the main root, number of primary and secondary roots, mycorrhizal colonization) and accumulation of nitrogen, phosphorus and potassium were measured by vector graphic analysis. The results obtained showed that there was a differentiated effect of mycorrhizae on the growth of S. mahagoni. Inoculation with mycorrhizal strains improved plant nutrition. The best substrate for the establishment of the mycorrhizal association was S2, composed of 20 % coconut husk + 60 % coconut fiber + 20 % pine sawdust, but the best treatments for the growth and nutrition of the seedlings were the combination of substrate S2 and the strains Glomus cubense and Rhizophagus irregulare.
Keywords: Vector analysis; Mycorrhizal colonization; Forest nutrition; Root system.
RESUMO
O estudo foi desenvolvido com o objetivo de avaliar a influência de fatores de substrato e cepas de fungos micorrízicos arbusculares (FMA) no sistema radicular e estado nutricional da Swietenia mahagoni L. Jacq. Os fatores testados foram: substrato fator 1: casca de cacau, fibra de coco e serrim de pinheiro compostado em proporções 6:2:2 e 2:6:2, e um controle composto de 100% de solo; cepas micorrízicas fator 2: Glomus cubense, Rhizoglomus irregulare e Funneliformis mosseae. Os nove tratamentos resultantes do arranjo fatorial 3x3 foram estabelecidos sob um desenho experimental completamente aleatório. Variáveis relacionadas com o sistema radicular (comprimento da raiz principal, número de raízes primárias e secundárias, colonização micorrízica) e acumulação de azoto, fósforo e potássio foram medidas por análise gráfica vetorial. Os resultados obtidos mostraram que houve um efeito diferenciado de micorrizas sobre o crescimento de S. mahagoni. A inoculação com as estirpes micorrízicas melhorou a nutrição vegetal. O melhor substrato para o estabelecimento da associação micorrizal foi o S2 constituído por 20 % de casca de coco + 60 % de fibra de coco + 20 % de serradura de pinheiro, mas os melhores tratamentos para o crescimento e nutrição das plântulas foram a combinação do substrato S2 e as estirpes Glomus cubense e Rhizophagus irregulare.
Palavras-chave: Análise vectorial; colonização micorrízica; nutrição florestal; sistema radicular.
INTRODUCCIÓN
Swietenia mahagoni L. Jacq. (caoba antillana) es probablemente una de las especies maderables más valiosas del mundo, la más conocida y apreciable para ebanistería y toda clase de obra que requiera madera de alta calidad. Es empleada para la reforestación en diversas áreas de Cuba por ser autóctona, heliófila facultativa, colonizadora de diferentes etapas sucesionales, tolerantes a la competencia, con amplia distribución en el país, posee diferentes estrategias de crecimiento y desarrollo y se reproduce fácilmente en vivero (Ricardo et al., 2016).
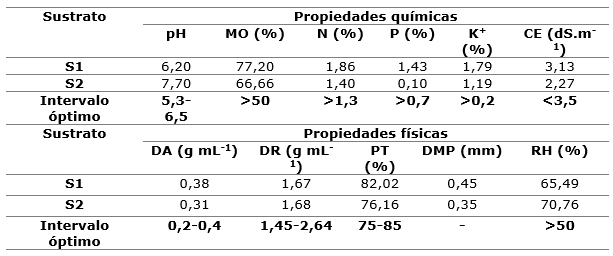
Para la producción en vivero es importante elegir un sustrato con característica química y física adecuada, que proporcione suficiente espacio para que las raíces crezcan, ya que de ella depende la calidad inicial y el buen crecimiento de las plántulas (Arévalo et al., 2016). Dichas características incluyen pH ligeramente ácido, fertilidad, libre de plagas y enfermedades, además de presentar valores mínimos en la porosidad total de 70 %, porosidad de aireación de 10 % y porosidad de retención de agua de 55 % (Abad et al., 2005).
Además de la elección adecuada del sustrato, una alternativa para obtener cantidad, calidad y mayor crecimiento de las plántulas es favorecer la formación de hongos micorrízicos arbusculares (HMA), cuya inoculación es particularmente apropiada en condiciones de viveros. Estos permiten a la planta ampliar la exploración del sustrato a través del micelio y transportar nutrientes a la raíz, así como incrementar la superficie de absorción de la planta (Oliveira Júnior et al., 2019), favoreciendo la absorción de los diferentes nutrimentos incluyendo los de baja movilidad en el suelo (Piliarová et al., 2019).
Sin embargo, la expresión benéfica de la simbiosis depende del tipo de sustrato, ya que su naturaleza y propiedades físicas y químicas afectan el establecimiento y comportamiento de los HMA. Tomando en consideración lo señalado anteriormente este estudio tiene como objetivo evaluar la influencia de diferentes sustratos y cepas de hongos micorrízicos arbusculares (HMA) sobre la morfología y estado nutricional de S. mahagoni.
MATERIALES Y MÉTODOS
Descripción del experimento
Se realizó un experimento en el vivero del Centro de Estudio de Tecnología Agroforestal (CETAF) perteneciente a la Universidad de Guantánamo (UG), localizado en las coordenadas geográficas 20°12´21´´de latitud Norte y los 75°13´37´´ de longitud Oeste a 87 msnm. El clima presenta una marcada estacionalidad seca de enero a marzo y de noviembre a diciembre; y lluvias desde abril hasta junio y septiembre hasta octubre. Este sitio posee una precipitación y temperatura media anual de 1 028 mm y 25,9 °C respectivamente (INSMET, 2020).
Las semillas de Swietenia mahagoni fueron obtenidas de frutos maduros recolectados de una masa ubicada en el municipio Jamaica perteneciente a la Empresa Agroforestal Guantánamo. Los frutos fueron procesados teniendo en cuenta los aspectos que establece para el beneficio de los mismos la Norma Cubana 318/1978. Las semillas se utilizaron inmediatamente después de su procesamiento. Previo a la siembra se realizó un ensayo de germinación con resultado de más de 90 % de semillas germinadas en todos los tratamientos.
El cultivo se realizó en contenedores plásticos tipo tubetes con una capacidad de 200 cm3. Después de la siembra y hasta el primer mes el riego se efectuó de forma manual, dos veces al día, por la mañana y por la tarde. A partir del segundo mes solo se efectuó un riego diario y un mes antes de finalizar el cultivo se comenzó el proceso de endurecimiento de las plantas consistentes en riego en días alternos.
Diseño experimental
El diseño experimental fue completamente al azar, con arreglo factorial y cinco replicas. Los factores fueron: A) cepas de HMA y B) sustratos. Adicionalmente se empleó un testigo compuesto por 90 % de suelo agrícola + 10 % de cascarilla de cacao (Instructivo técnico), para un total de nueve tratamientos (Tabla 1).
Tabla 1. - Descripción de los tratamientos aplicados en el experimento
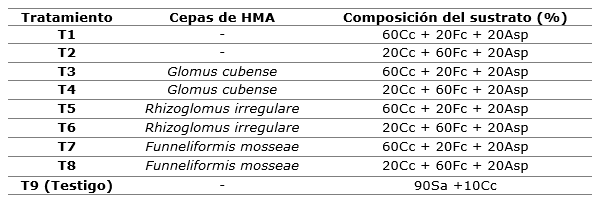
Sa: suelo agrícola; Cc: cascarilla de cacao; Fc: fibra de coco; Asp: aserrín de pino.
Las cepas HMA utilizadas fueron: Funneliformis mosseae, Glomus cubense y Rhizoglomus irregulare, con 30, 33 y 36 esporas g-1 de inoculante respectivamente, procedentes del cepario del Instituto Nacional de Ciencias Agrícolas (INCA). La inoculación se realizó por el método de recubrimiento de la semilla (Fernández et al., 2001).
Los sustratos se esterilizaron a 120°C durante 15 minutos en autoclave por tres ocasiones, y estos fueron: S1) cascarilla de cacao 60 % + fibra de coco 20 % +aserrín de pino 20 % (6:2:2) y S2) 20 % de cascarilla de caco + 60 % de fibra de coco + 20 % de aserrín de pino (6:2:2). En la Tabla 2, se presentan las características químicas y físicas de los sustratos y sus intervalos óptimos sugeridos por Abad et al., (2005) y Arévalo et al., (2016), los que se encuentran entre los valores óptimos, no así para el sustrato S2 en el que el fósforo se encuentra por debajo del intervalo óptimo (Tabla 2).
Tabla 2. - Características químicas y físicas de los sustratos
Variables evaluadas
Las variables evaluadas al final del experimento (120 días) incluyeron: largo de la raíz principal (cm), medida desde el cuello hasta el ápice, mediante el empleo de una regla graduada; la cantidad de raíces finas y gruesas (No.) fueron determinadas mediante conteo visual. La colonización por HMA en raíz, se obtuvo a partir de la tinción de las raíces según la metodología descrita por Yon et al., (2015) y posteriormente se determinó el porcentaje de colonización micorrízica con el método descrito por Mcgonigle et al., (1990). Se tomaron muestras foliares para determinar: concentraciones de nitrógeno (%) mediante el método de Kjeldahl, fósforo (%) por el colorimétrico y el potasio (%) por flamometría con un fotómetro de llama.
Análisis estadístico
Los datos obtenidos en las variables evaluadas se sometieron a análisis de varianza y comparación de medias mediante la prueba de Tukey (α=0,05 %). Se evaluó en grado de asociación entre las variables mediante un análisis de correlación a través del coeficiente de Pearson. Asimismo, se realizó un análisis de componentes principales para todos los indicadores evaluados. Todo ello utilizando el programa estadístico SPSS ver. 23 para Windows.
El estado nutricional de las plántulas se diagnosticó por el método gráfico de vectores propuesta por Park et al., (2015), el cual analiza, conjuntamente, los cambios relativos de las variables masa del componente vegetal, concentración y contenido de nutrimentos. Los vectores se graficaron con un programa basado en macros programadas en Excel (http://bit.ly/2Eok8GF). Los efectos principales se determinaron de acuerdo con la interpretación gráfica, tal como lo sugieren López y Alvarado (2010) y Xiao et al., (2019).
RESULTADOS Y DISCUSIÓN
Aunque se observó una respuesta significativa en todas las variables evaluadas en S. mahagoni al final del cultivo con la aplicación conjunta de sustrato y micorriza, fue el factor micorriza el que generó la mayor variación en estos parámetros, superando los niveles de variabilidad generados por el factor sustrato y la interacción entre ambos (Tabla 3). Esto sugiere que independientemente de las concentraciones del sustrato, los hongos micorrízicos son capaces de facilitar el crecimiento vegetal. Los resultados coinciden con estudios realizados en otras especies, donde se destacaron los tratamientos micorrizados (Aguirre-Medina et al., 2019; Oliveira Júnior et al., 2019; Quiñones-Aguilar et al., 2020).
Tabla 3. - Cuadrados medios y nivel de significancia de parámetros del sistema radical y nutricional de S. mahagoni al final del cultivo
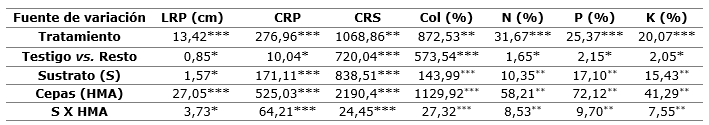
*= P< 0,05; **= P< 0,01; ***= P< 0,001; LRP= largo de la raíz principal; CRP y
CRS= cantidad de raíces primarias y secundarias; Col= porcentaje de colonización micorrízica;
N= nitrógeno; P= fósforo; P= potasio; HMA= hongos micorrízicos arbusculares.
Morfología del sistema radical y colonización micorrízica
En relación al análisis de los atributos relacionados con el sistema radical (Tabla 4) se observó que las plántulas se ven beneficiadas con la combinación del sustrato S2 conformado por 20 % de cascarilla de cacao + 60 % de fibra de coco + 20 % aserrín de pino en interacción con las cepas micorrízicas G. cubense y R. irregulare (T4 y T6), con incremento con respecto al testigo de 15 y 10 % para LRP, 25 y 20 % para CRP, 30 y 25 % para CRS. Los menores valores correspondieron a los tratamientos no micorrizados, lo que corrobora que la micorriza beneficia la emisión de raíces (Aguirre-Medina et al., 2019), en correspondencia con valores altos de porosidad y buena aireación, lo que favorece el crecimiento de raíces y, por ende, el desarrollo de la parte aérea de la planta.
En este sentido, algunos autores han señalado que las micorrizas son hongos beneficiosos, que pueden incrementar el desarrollo de las raíces hasta 200 % en un grupo de plantas, siendo un importante componente de la microflora del suelo, señalándose que pueden aumentar la absorción de nutrimentos por las raíces entre siete y 250 veces dependiendo del cultivo (Alvero et al., 2011).
Tabla 4. - Efecto de los sustratos orgánicos y la micorriza arbuscular en el sistema radical de S. mahagoni al final del cultivo
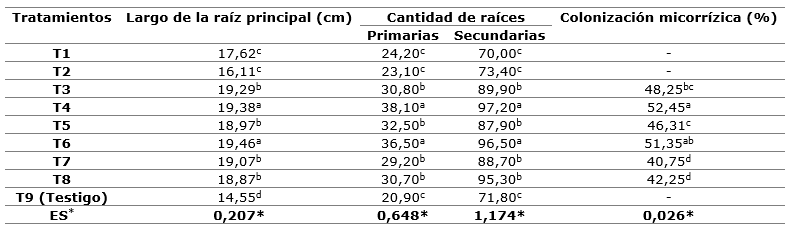
Letras diferentes en una misma columna difieren para P <0,05.ES- Error estándar.
La mayor cantidad de raíces secundarias obtenido en los tratamientos T4 y T6, pudiera estar dada por las características físicas del sustrato (Tabla 2), en el cual se generó un número mayor de raíces secundarias, capaces de colonizar más rápido el cepellón. Al respecto, Álvaro et al., (2014) citado por López et al., (2020) afirman que la abundante emisión de raíces secundarias demuestra alta calidad y garantiza un rápido crecimiento de las plantas después de la plantación; además hacen referencia también que el número de raíces secundarias de primer orden han mostrado correlación para mejorar el desempeño de las plantas en el campo.
En el caso de los sustratos inoculados, la diferencia está marcada a favor del sustrato S2, beneficios que pueden estar relacionados con el bajo contenido de fósforo (Tabla 2). En este caso, la actividad fúngica asociada a la planta genera un sistema de beneficio mutuo, pues los HMA modifican la morfología de las raicillas, dándole a la planta una mejor capacidad para alcanzar el agua y disolver los nutrientes, pudiendo resolver el problema del agotamiento inminente del stock de fosfato y otros elementos como, nitrógeno, zinc, cobre, hierro, potasio, calcio y magnesio. También puede causar un cambio en la absorción de nutrientes y al mismo tiempo permiten la inducción de las defensas del hospedero (Uc-Ku et al., 2019; Piliarová et al., 2019).
Por su parte, Brito et al. (2017) indican que la actividad y el beneficio de la simbiosis son más visibles cuando estos se encuentran en suelos deficientes en fósforo. En esta condición, plantas de Schizolobium parahyba var. amazonicum (Huber ex Ducke) Barneby inoculadas con el hongo Rhizophagus clarus presentaron mayores tasas de crecimiento con relación al control. Esta situación podría explicar el comportamiento de las cepas estudiadas en este experimento, pues el sustrato S1 se caracterizó por valores superiores de fósforo disponible.
De manera general, los sustratos evaluados (S1 y S2) resultaron superiores al testigo de producción donde el suelo es utilizado en mayor proporción, el cual resultó la de menor respuesta entre los sustratos evaluados para esta etapa o fase de desarrollo del cultivo, por lo que sería recomendable su utilización solo en el caso de no contar con otros materiales complementarios, teniendo en cuenta además lo señalado en la literatura, de no usar tierra en contenedores con volúmenes pequeños porque se pueden presentar problemas fitosanitarios y mal desarrollo radical (Valenzuela, 2019).
El mayor crecimiento radical de plantas de caoba obtenidas en sustratos orgánicos inoculados con diferentes especies y combinaciones de HMA ya ha sido reportada previamente (Gómez et al., 2018; Rajan et al., 2020; Falcón et al., 2020). Estos autores expresan que la efectividad en el crecimiento posiblemente se deba a una influencia sinérgica al combinar HMA y sustratos orgánicos. Por otro lado, la variabilidad en el crecimiento de las plantas de caoba asociadas con hongos micorrízicos puede deberse varios factores como la especie de hongo micorrízico, la especie de caoba, la complementariedad funcional o sinergismo de los hongos micorrízicos y, a la respuesta de cada hospedero (alta o baja dependencia a los hongos micorrízicos) (Abd El-Kader et al., 2016).
La colonización micorrízica de la raíz mostró diferencias estadísticas significativas (Tukey p≤0,05) por efecto de los tratamientos de HMA (Tabla 4). Los mayores porcentajes de micorrización se obtuvieron en el sustrato S2 inoculados con G. cubense y R. irregulare (T4 y T6) con valores de 52,45 y 51,55 %, respectivamente, sin diferencias estadística con respecto al sustrato S1 inoculados con las mismas cepas (T3 y T5). Estos valores de micorrización fueron significativamente superiores a los alcanzados con F. mosseae en ambos sustratos (40,75 y 42,25 %). En todos los sustratos inoculados se presentaron porcentajes de micorrización entre 40,75 y 52,45 %. Esto permite afirmar que hay cierto grado de dependencia entre la planta y el HMA, elemento de gran importancia debido a que, en la simbiosis micorrízica, el hongo permite el incremento en la adquisición de nutrientes, principalmente de aquellos de escasa movilidad en el suelo, como el fósforo, al mismo tiempo que la planta proporciona compuestos carbonados para el crecimiento del hongo (Mei et al., 2019).Estos resultados muestran la especificidad infectiva que establecen los HMA en las plantas, dependiendo de las características de cada sustrato (Feijen et al., 2018).
Resultados similares fueron descritos por Aguirre-Medina et al., (2019), quienes inocularon HMA en Tabebuia donnell-smithii Rose, encontrando altos niveles de micorrización. En otros estudios se ha observado que las plantas de caoba antillana son colonizadas por Glomus hoi like (Oconor et al., 2011), lo que es comparable con los resultados encontrados en el presente trabajo, tanto con la cepa G. cubense como R. irregulare, puesto que ambas colonizaron las raíces de caoba antillana, destacándosela cepa G. cubense.
En la Tabla 5, se muestran los resultados del análisis de correlación, en ella se puede evidenciar que el porcentaje de colonización micorrízica presentó una correlación positiva y significativa con todas las características químicas del suelo evaluadas, excepto con el contenido de materia orgánica y diámetro medio de las partículas. El largo de la raíz principal, cantidad de raíces primarias y secundarias presentaron correlaciones positivas y significativas con la porosidad total.
Este resultado puede indicar que el porciento de colonización tuvo una estrecha relación con algunas propiedades del sustrato. Al respecto, se ha reportado que, en presencia de una alta disponibilidad de nutrientes, la respuesta de las plantas a la inoculación micorrízica se ve limitada, aunque esta respuesta va a depender de varios factores, entre ellos, el pH que puede afectar la absorción de nutrientes (fundamentalmente P) por la planta (Martín-Alonso et al., 2017).
En este trabajo, la disponibilidad de nutrientes del sustrato no limitó el porciento de colonización, por el contrario, influyó en el grado de colonización de las raíces. No obstante, Ji y Bever (2016), refieren que plantas que crecen en bajas condiciones de fósforo, como las desarrolladas en el sustrato S2, incrementan exudación de aminoácidos, reduciendo los azúcares y ácidos carboxílicos, lo que favorece a la micorrización. Además de lo expresado, la porosidad total, capacidad de retención de agua y capacidad de aireación fueron óptimos en ambos sustratos los que favorecieron la colonización (Tabla 5).
Tabla 5. - Coeficiente de correlación de Pearson entre las características químicas-físicas del sustrato y las variables del sistema radical evaluadas en S. mahagoni
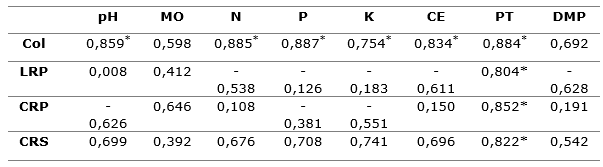
*Correlaciones significativamente diferentes de cero, con un nivel de confianza del 95,0 %;
LRP: largo de la raíz principal; CRP y CRS: cantidad de raíces primarias y secundarias;
Col:
porcentaje de colonización micorrízica; PT: porosidad total; DMP: diámetro medio de las partículas;
MO: materia orgánica.
En el caso del largo de la raíz principal, cantidad de raíces primarias y secundarias es posible que la correlación positiva con la porosidad total se deba al porciento de esta, el cual se encuentra por encima de 75 % en ambos sustratos, característica que facilita la infiltración, profundidad radical efectiva y circulación de aire, estimulando el desarrollo uniforme y abundante del sistema radical, lo cual favorece el crecimiento y desarrollo de las plantas (Abad et al., 2005).
En el análisis de componentes principales para las características químicas-físicas de los sustratos con las variables evaluadas (Figura 1), se observó la formación de dos componentes, los que permitieron explicar el 86,35 % de la variabilidad total, siendo el 61,18 % de la varianza explicada por la componente principal 1 (eje horizontal).
Las variables que más contribuyeron a la formación de la primera componente fueron el porcentaje de colonización micorrízica, cantidad de raíces secundarias porosidad total, conductividad eléctrica, pH del sustrato y los contenidos de N, P, K y DMP. Para la segunda componente, las variables con mayor contribución fueron el largo de la raíz principal, cantidad de raíces primarias y el porcentaje de materia orgánica del sustrato.
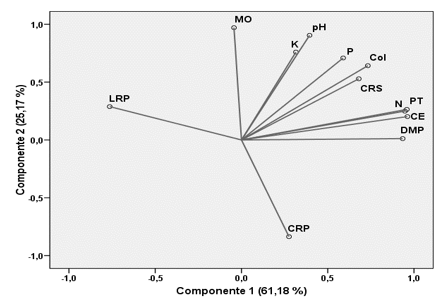
Figura 1. - Resultados del Análisis de Componentes Principales
LRP: largo de la raíz principal; CRP y CRS: cantidad de raíces primarias y secundarias;
Col: porcentaje de colonización micorrízica;
PT: porosidad total; DMP: diámetro medio de las
partículas; MO: materia orgánica.
En la Figura 1, se observa como el pH del sustrato tiene una estrecha relación con las variables relacionadas con la disponibilidad de nutrientes, que se agrupan en el cuadrante superior derecho, este resultado puede que indique que es el pH el principal factor que va a determinar el grado de disponibilidad de los nutrientes, coincidiendo con Martín-Alonso et al., (2017), quienes señalan que la disponibilidad de los nutrientes tienen una alta relación con el pH lo cual determinó la respuesta de Canavalia ensiformis ante la inoculación micorrízica.
Al respecto, Kawahara et al., (2016) plantean que la relación que se establece entre los rangos de pH del sustrato y el efecto de la colonización micorrízica es verdaderamente complejo, dependiendo no sólo de la especie micótica, sino también del tipo de suelo, la forma en que se encuentran los nutrientes (fundamentalmente P y N y otros elementos como Cu, Zn, Mo, B, etc.) y en menor medida de la especie de planta sobre la que se desarrolla.
Análisis nutricional
Independientemente del efecto correspondiente entre los factores evaluados, las plantas con mayor cantidad de reservas nutricionales fueron las producidas en los tratamientos inoculados. La Figura 2 muestra la tendencia de vectores para biomasa aérea relativa (cambio en biomasa aérea con respecto al testigo) de este estudio. De acuerdo con los nomogramas de vectores y al tomar como punto de referencia el estado nutricional de las plantas del tratamiento testigo (T9), en nitrógeno las plantas de los tratamientos no micorrizados (T1 y T2) tuvieron un efecto de dilución y las de los tratamientos micorrizados (T3, T5, T7 y T8) y (T4 y T6), evidenciaron suficiencia y consumo de lujo, respectivamente(Figura 2A), lo cual indica incremento en la absorción de este elemento a través de la formación simbiótica de órganos en la raíz (Feijen et al., 2018), permitiendo así que las plantas sean más resistentes a diferentes cambios adversos que puedan existir en un ecosistema.
Alvarado-Hernández y Raigosa (2012), subrayan que el nitrógeno es un constituyente de las proteínas, la clorofila, los ácidos nucleicos y otras sustancias de las plantas. Un suministro adecuado de nitrógeno promueve el crecimiento vegetal, incrementa la relación biomasa/raíces, y es esencial para la formación de tallos, ramas y hojas.
Por otra parte, se ha demostrado que los HMA participan activamente en la transferencia de N del suelo a la planta hospedera (Bücking y Kafle, 2015). A partir de estas premisas, se podía esperar un efecto positivo de la inoculación con R. irregulare en la absorción de N, en el incremento en las concentraciones de este elemento en la biomasa aérea, como se observó en la Figura 2A y, en consecuencia, una mejora de la nutrición nitrogenada de la especie.
El aporte de fósforo (Figura 2B) fue de lujo en los tratamientos micorrizados (T3, T4, T5, T6, T7 y T8), no así en los tratamientos no micorrizados (T1 y T2) las que presentaron una dilución en el tejido foliar, por lo que no aportaron las cantidades suficientes. López y Alvarado (2010) plantean que bajo estas condiciones el elemento en estudio puede limitar el crecimiento. El consumo de lujo en las plantas micorrizadas se denota por el desplazamiento mayor de cada vector hacia la esquina superior derecha de la figura 2B. En ese contexto los resultados obtenidos en la absorción de fósforo en este estudio, demuestran que los HMA jugaron un papel muy importante en la traslocación de este nutriente a la especie forestal evaluada, ya que las hifas del hongo actúan como una extensión de la raíz de la planta, permitiendo que la longitud absorbente de la raíz crezca y por consiguiente la exploración del suelo también aumente. Las raíces pueden llegar hasta donde el fósforo se encuentra y así facilitar la absorción del mismo (Mei et al., 2019). Además, se ha observado que la asociación micorrízica aumenta el contenido de fósforo en las hojas de Swietenia mahagoni (Falcón et al., 2020).
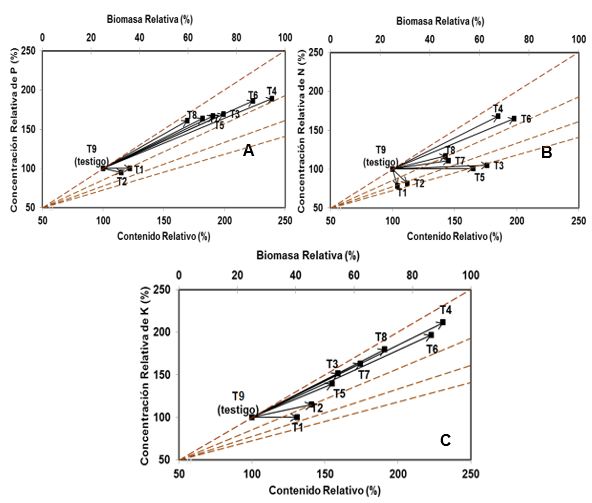
Figura 2. - Nomograma de vectores del estado nutricional de nitrógeno (N), fósforo (P) y potasio (K) de plántulas de S. mahagoni al final del cultivo
Con respecto al potasio, las plantas de los tratamientos micorrizados (T3, T4, T5, T6, T7 y T8) tuvieron en común efectos de consumo de lujo; no obstante, esta respuesta fue más representativa en los tratamientos T4 y T6. En las plántulas de los tratamientos T1 y T2 se registró un efecto de suficiencia, por lo que este elemento no limita el incremento del peso seco (Figura 2C). Según Alvarado-Hernández y Raigosa (2012), el potasio, después del nitrógeno es el elemento esencial requerido en mayores cantidades por la planta. Los mismos autores sostienen que la mayoría de especies vegetales tienen preferencia por la absorción de este nutrimento, puesto que ejerce un papel como transportador de otros elementos, además de que es esencial para la osmorregulación y el control estomático.
Resulta de interés destacar que la adición de los distintos HMA hayan sido efectivo para mejorar el estado nutricional en la biomasa de la parte aérea y el crecimiento de S. mahagoni, a pesar de las concentraciones de fósforo en los sustratos, pues algunos trabajos refieren que el efecto de la inoculación micorrízica puede inhibirse en presencia de altos contenidos de este elemento (Ji y Bever, 2016). Sin embargo, otros han encontrado que las plantas inoculadas con cepas de HMA eficientes, pueden alcanzar un funcionamiento micorrízico efectivo, aun en sustratos con altas concentraciones de fósforo, siempre que otros nutrientes limiten su crecimiento y desarrollo (Mei et al., 2016).
Resultados similares encontraron Gómez et al., (2018), al investigar la variación de la eficiencia nutricional y el crecimiento de cuatro especies forestales (Pachira quinata (Jac.) W.S. Alverson, Gmelina arborea Roxb, Eucalyptus sp. y Acacia mangium Willd.), por la infección de cuatro inóculos de HMA, donde la eficiencia de la absorción de los macro y micronutrientes fue mayor en las plantas inoculadas que en las no inoculadas, especialmente en los suelos con bajo fósforo disponible.
Los resultados obtenidos al final del cultivo en vivero, demostraron que el mejor sustrato para el establecimiento de la asociación micorrízica fue el S2 (20 % cascarilla de caco + 60 % de fibra de coco + 20 % de aserrín de pino), pero el mejor tratamiento para el crecimiento radical de las plantas fue con este sustrato y con las cepas Glomus cubense y Rhizophagus irregulare. La inoculación con las cepas micorrízicas mejoró el estado nutricional de las plantas.
Estos resultados pueden apoyar la toma de decisiones en las actividades de reforestación relacionadas con la especie S. mahagoni y su cultivo en viveros con la tecnología de tubetes.
REFERENCIAS BIBLIOGRÁFICAS
ABAD, M., FORNES, F., CARRIÓN, C., NOGUERA, V., NOGUERA, P., MAQUIEIRA, A. y PUCHADES, R., 2005. Physical Properties of Various Coconut Coir Dusts Compared to Peat. HortScience [en línea], vol. 40, no. 7. DOI 10.21273/HORTSCI.40.7.2138. Disponible en: https://www.researchgate.net/publication /268430441_Physical_Properties_of_Various_Coconut_Coir_Dusts_Compared_to_Peat .
ABD EL-KADER, M.M.E., NAGLAA,K., A.-E. y ABO-ELSOUD, I.H., 2016. Effects of Sewage Sludge and Endo- mycorrhizal on Growth, Chemical Content and Some Physical Properties of Swietenia mahagoni seedling. Annals of Agricultural Science, Moshtohor [en línea], vol. 54, no. 4, pp. 905-918. [Consulta: 31 mayo 2021]. ISSN 1110-0419. DOI 10.21608/assjm.2016.112675. Disponible en: https://assjm.journals.ekb.eg/article_112675.html.
AGUIRRE-MEDINA, J.F., YEEKÓN-MÉNDEZ, L. y ESPINOSA-ZARAGOZA, S., 2019. Influencia de hongos endomicorrízicos en el crecimiento de (Tabebuia donnell-smithii Rose). Ecosistemas y recursos agropecuarios [en línea], vol. 6, no. 16, pp. 11-21. [Consulta: 31 mayo 2021]. ISSN 2007-9028. DOI 10.19136/era.a6n16.1538. Disponible en: http://www.scielo.org.mx/scielo.php?script=sci_abstract&pid=S2007-90282019000100011&lng=es&nrm=iso&tlng=es .
ALVARADO-HERNANDEZ, A. y RAIGOSA, J., 2012. Nutrición y Fertilización Forestal en Regiones Tropicales [en línea]. 1ra ed: San José, Costa Rica: Asociación Costarricense de la Ciencia del Suelo. ISBN 978-9968-9422-5-6. Disponible en: https://www.researchgate.net/publication/324165376_Nutricion_y_Fertilizacion_Forestal_en_Regiones_Tropicales .
ALVERO, C.M., NARANJO, J.M.C., GOROTIZA, J.C. y ESPINOSA, R.R., 2011. Evaluación de sustratos y aplicación de hongos micorrízicos arbusculares en Begonia sp. Cultivos Tropicales [en línea], vol. 32, no. 2, pp. 17-22. [Consulta: 31 mayo 2021]. ISSN 1819-4087. Disponible en: https://www.redalyc.org/articulo.oa?id=193222422001 .
ARÉVALO P, M.E., OBERPAUR W, C. y MÉNDEZ C, C., 2016. Inclusión de musgo (Sphagnum magellanicum Brid.) y fibra de coco como componentes orgánicos del sustrato para almácigos de kiwi (Actinidia deliciosa). Idesia (Arica) [en línea], vol. 34, no. 2, pp. 47-55. [Consulta: 31 mayo 2021]. ISSN 0718-3429. DOI 10.4067/S0718-34292016005000007. Disponible en: http://www.scielo.cl/scielo.php?script=sci_abstract&pid=S0718-34292016000200007&lng=es&nrm=iso&tlng=es .
BRITO, V.N., TELLECHEA, F.R.F., HEITOR, L.C., FREITAS, M.S.M. y MARTINS, M.A., 2017. Fungos micorrízicos arbusculares e adubação fosfatada na produção de mudas de paricá. Ciência Florestal [en línea], vol. 27, pp. 485-497. [Consulta: 31 mayo 2021]. ISSN 1980-5098, 0103-9954, 1980-5098. DOI 10.5902/1980509827730. Disponible en: https://www.scielo.br/j/cflo/a/wtgMFtwr7kNCZmhCDYT7ZJt/abstract/?lang=pt .
BÜCKING, H. y KAFLE, A. 2015. Role of arbuscular mycorrhizal fungi in the nitrogen uptake of plants: current knowledge and research gaps. Agronomy [en línea], vol. 5, no. 4, pp. 587612. [Consulta: 31 mayo 2021]. ISSN 2073-4393. DOI 10.3390/agronomy5040587. Disponible en: https://www.mdpi.com/2073-4395/5/4/587 .
FALCÓN, E., COBAS, M. y BONILLA, M., 2020. Influencia de hongos micorrizógenos arbusculares en el crecimiento y nutrición de Swietenia mahagoni L. Jacq. Hombre, Ciencia y Tecnología [en línea], vol. 24, no. Número especial, pp. 25-32. Disponible en: https://www.ciencia.gtmo.inf.cu/index.php/http/article/view/1089 .
FEIJEN, F.A.A., VOS, R.A., NUYTINCK, J. y MERCKX, V.S.F.T., 2018. Evolutionary dynamics of mycorrhizal symbiosis in land plant diversification. Scientific Reports [en línea], vol. 8, no. 1, pp. 10698. [Consulta: 31 mayo 2021]. ISSN 2045-2322. DOI 10.1038/s41598-018-28920-x. Disponible en: https://www.nature.com/articles/s41598-018-28920-x .
FERNÁNDEZ, F., GÓMEZ, R., VANEGAS, L., NOVAL, B.M. y MARTINEZ, M.A., 2001. Producto inoculante micorrizógeno. 2001. S.l.: Oficina Nacional de Propiedad Industrial. Cuba, Patente, 22641.
GÓMEZ, M.M.R., ROLON, A.M.P., MONCADA, U.A.P. y SERRALDE, D.P., 2018. Biofertilización con hongos formadores de micorrizas arbusculares (HFMA) en especies forestales en vivero. Biotecnología en el Sector Agropecuario y Agroindustrial [en línea], vol. 16, no. 2, pp. 15-25. [Consulta: 31 mayo 2021]. ISSN 1909-9959. DOI 10.18684/bsaa.v16n2.1162. Disponible en: https://revistas.unicauca.edu.co/index.php/biotecnologia/article/view/1162 .
INSTITUTO DE METEOROLOGÍA DE CUBA [INSMET], 2020. Centro Meteorológico Provincial de Guantánamo [en línea]. 2020. S.l.: Instituto de Meteorología de Cuba [INSMET]. Disponible en: http://www.guantanamo.gob.cu/es/centro-meteorologico-provincial-guantanamo.
JI, B. y BEVER, J.D., 2016. Plant preferential allocation and fungal reward decline with soil phosphorus: implications for mycorrhizal mutualism. Ecosphere [en línea], vol. 7, no. 5, pp. e01256. [Consulta: 31 mayo 2021]. ISSN 2150-8925. DOI https://doi.org/10.1002/ecs2.1256. Disponible en: https://esajournals.onlinelibrary.wiley.com/doi/abs/10.1002/ecs2.1256 .
KAWAHARA, A., AN, G.-H., MIYAKAWA, S., SONODA, J. y EZAWA, T., 2016. Nestedness in arbuscular mycorrhizal fungal communities along soil pH gradients in early primary succession: Acid-tolerant fungi are pH generalists. PLOS ONE [en línea], vol. 11. DOI 10.1371/journal.pone.0165035. Disponible en: https://www.researchgate.net/publication /309281188_Nestedness_in_Arbuscular_Mycorrhizal_Fungal_Communities_along_Soil_pH_Gradients_in_Early_Primary_Succession_Acid -Tolerant_Fungi_Are_pH_Generalists.
LÓPEZ, M.C., SOSPEDRA, R.S. y RAMOS, Y.A., 2020. Comportamiento de los parámetros morfológicos de calidad de la planta de Lysiloma sabicu Benth. en vivero sobre sustratos orgánicos. Revista Cubana de Ciencias Forestales [en línea], vol. 8, no. 3, pp. 550-561. [Consulta: 31 mayo 2021]. ISSN 2310-3469. Disponible en: https://cfores.upr.edu.cu/index.php/cfores/article/view/615 .
LÓPEZ-LÓPEZ, M.Á. y ALVARADO-LÓPEZ, J., 2010. Interpretación de nomogramas de análisis de vectores para diagnóstico nutrimental de especies forestales. Madera y bosques [en línea], vol. 16, no. 1, pp. 99-108. [Consulta: 31 mayo 2021]. ISSN 1405-0471. Disponible en: http://www.scielo.org.mx/scielo.php?script=sci_abstract&pid=S1405 -04712010000100007&lng=es&nrm=iso&tlng=es .
MARTÍN-ALONSO, G.M., TAMAYO-AGUILAR, Y., RAMÍREZ-PEDROSO, J.F., VARELA-NUALLES, M., RIVERA-ESPINOSA, R., 2017. Relación entre la respuesta de Canavalia ensiformis a la inoculación micorrízica y algunas propiedades químicas del suelo. Revista Cultivos Tropicales [en línea], vol. 38, no. 3, pp. 24-29. [Consulta: 31 mayo 2021]. ISSN 1819-4087. Disponible en: http://www.redalyc.org/articulo.oa?id=193253129003 .
MCGONIGLE, T.P., MILLER, M.H., EVANS, D.G., FAIRCHILD, G.L. y SWAN, J.A., 1990. A new method which gives an objective measure of colonization of roots by vesiculararbuscular mycorrhizal fungi. New Phytologist [en línea], vol. 115, no. 3, pp. 495-501. [Consulta: 31 mayo 2021]. ISSN 1469-8137. DOI https://doi.org/10.1111/j.1469-8137.1990.tb00476.x. Disponible en: https://nph.onlinelibrary.wiley.com/doi/abs/10.1111/j.1469-8137.1990.tb00476.x.
MEI, L., YANG, X., ZHANG, S., ZHANG, T. y GUO, J., 2019. Arbuscular mycorrhizal fungi alleviate phosphorus limitation by reducing plant N: P ratios under warming and nitrogen addition in a temperate meadow ecosystem. Science of The Total Environment [en línea], vol. 686, pp. 1129-1139. [Consulta: 31 mayo 2021]. ISSN 0048-9697. DOI 10.1016/j.scitotenv.2019.06.035. Disponible en: https://www.sciencedirect.com/science/article/pii/S0048969719325951.
OCONOR, E., NELSON, M.R. y LEYVA, O., 2011. Efecto de la aplicación de micorrizas arbusculares sobre la producción de posturas de Caoba antillana (Swietenia mahagoni L. Jacq. Revista Electrónica Hombre, Ciencia y Tecnología [en línea], vol. 14, no. 4, pp. 1-8. [Consulta: 31 mayo 2021]. Disponible en: https://www.ciencia.gtmo.inf.cu/index.php/http/article/view/177.
OLIVEIRA JÚNIOR, J.Q., RODRIGUES DA SILVA, A.C., MOREIRA DA SILVEIRA, P.V., CONCEIÇÃO E.J. y PEREIRA, M.G., 2019. Effect of mycorrhizal inoculation and substrate composition on seedling growth of two Atlantic Forest Tree Species. Floresta, Curitiba [en línea], vol. 49, no. 4, pp. 623-632. [Consulta: 31 mayo 2021]. ISNN 1982-4668. DOI 10.5380/rf.v49 i4.54525. Disponible en: https://doi.org/10.5380/rf.v49i4 .
PARK, B.B., PARK, G.E. y BAE, K., 2015. Diagnosis of plant nutrient and growth responses on fertilization with vector analysis and morphological index. Forest Science and Technology [en línea], vol. 11, no. 1, pp. 1-10. [Consulta: 31 mayo 2021]. ISSN 2158-0103. DOI 10.1080/21580103.2014.931257. Disponible en: https://doi.org/10.1080/21580103.2014.931257 .
PILIAROVÁ, M., ONDREICKOVA, K., HUDCOVICOVA, M., MIHÁLIK, D. y KRAIC, J., 2019. Arbuscular Mycorrhizal Fungi Their Life and Function in Ecosystem. Agriculture (Pol'nohospodárstvo) [en línea], vol. 65, no. 1, pp. 3-15. DOI 10.2478/agri-2019-0001. Disponible en: https://www.researchgate.net/publication/332981888_Arbuscular_Mycorrhizal_Fungi_-_Their_Life_and_Function_in_Ecosystem .
QUIÑONES-AGUILAR, E.E., RINCÓN-ENRÍQUEZ, G. y LÓPEZ-PÉREZ, L., 2020. Hongos micorrízicos nativos como promotores de crecimiento en plantas de guayaba (Psidium guajava L.). Terra Latinoamericana [en línea], vol. 38, no. 3, pp. 541-554. [Consulta: 31 mayo 2021]. ISSN 0187-5779. DOI 10.28940/terra.v38i3.646. Disponible en: http://www.scielo.org.mx/scielo.php?script=sci_abstract&pid=S0187-57792020000400541&lng=es&nrm=iso&tlng=es .
RAJAN, L., AV, S., K., S. y TK, K., 2020. Arbuscular Mycorrhizal Fungi Inoculation as a Climate Adaptation Strategy for Establishment of Swietenia macrophylla King. Seedlings. Forests [en línea], vol. 11, no. 488, pp. 2-15. DOI 10.3390/f11050488. Disponible en: https://www.researchgate.net/publication /340933416_Arbuscular_Mycorrhizal_Fungi_Inoculation_as_a_Climate_Adaptation_Strategy_for_Establishment_of_Swietenia_macrophylla_King_Seedlings .
RICARDO, N. E., PUENTES, D. A. y TORRES, Y. A., 2016. Relaciones entre las características morfológico-funcionales y ecológicas de especies autóctonas de meliáceas cubanas: estrategia para su empleo forestal. Acta Botánica Cubana [en línea], vol. 215, no. 1, pp. 2-23. [Consulta: 31 mayo 2021]. ISNN 2519-7724. Disponible en: https://www.revista.geotech.cu/index.php.abc .
UC-KU, A.G., ARREOLA-ENRÍQUEZ, J., CARRILLO-AVILA, E., OSNAYA-GONZÁLEZ, M.M., ALARCÓN, A., FERRERA-CERRATO, R., LANDEROS-SÁNCHEZ, C., UC-KU, A.G., ARREOLA-ENRÍQUEZ, J., CARRILLO-AVILA, E., OSNAYA-GONZÁLEZ, M.M., ALARCÓN, A., FERRERA-CERRATO, R. y LANDEROS-SÁNCHEZ, C., 2019. Inoculación de hongos micorrízicos arbusculares en el cultivo de Heliconia stricta. Revista mexicana de ciencias agrícolas [en línea], vol. 10, no. 5, pp. 1057-1069. [Consulta: 31 mayo 2021]. ISSN 2007-0934. DOI 10.29312/remexca.v10i5.1608. Disponible en: http://www.scielo.org.mx/scielo.php?script=sci_abstract&pid=S2007-09342019000501057&lng=es&nrm=iso&tlng=es .
VALENZUELA, O., 2019. El cultivo sin suelo y la sustentabilidad de las producciones intensivas. En: Accepted: 2019-04-09T12:32:44Z, Ediciones INTA (Boletín de divulgación técnica [en línea], no. 24, pp. 16. [Consulta: 31 mayo 2021]. ISSN 0327-3237. Disponible en: http://repositorio.inta.gob.ar:80/handle/20.500.12123/4853 .
XIAO, J., SUN, G., HAO, L., DONG, G. y ZHANG, Z., 2019. Afforestation and Reforestation: Drivers, Dynamics, and Impacts [en línea]. S.l.: MDPI. ISBN 978-3-03921-447-1. Disponible en: https://books.google.com.mx/books/about/Afforestation_and_Reforestation_Drivers.html?id=p9a2DwAAQBAJ&redir_esc=y .
YON, Y.R., PÉREZ, L.A., CARMONA, A.M., PÉREZ, Y.M., GARCÍA, L.R.M., SUÁREZ, K.F. y ECHEVARRÍA, A.M., 2015. Alternativa de la técnica de tinción para determinar la colonización micorrízica. Cultivos Tropicales [en línea], vol. 36, no. 2, pp. 18-21. [Consulta: 31 mayo 2021]. ISSN 1819-4087. Disponible en: https://www.redalyc.org/articulo.oa?id=193239249003 .
Conflicto de intereses:
Los autores declaran no tener conflictos de
intereses.
Contribución de los autores:
Los autores han participado en la redacción del trabajo y análisis de los documentos.
![]()
Esta obra está bajo una licencia de Creative Commons Reconocimiento-NoComercial
4.0 Internacional.
Copyright (c) 2021 Emir Falcón Oconor, Milagros Cobas López, Marta Bonilla Vichot, Orfelina Rodríguez Leyva