Revista Cubana de Ciencias Forestales. 2021; enero-abril 9(1): 53-71![]()
Efecto del estadio sucesional del bosque sobre la relación hospederos-orquídeas epífitas en la estación biológica Pindo Mirador, Ecuador
Effect of forest successional stage on the host-epiphyte orchid relationship in the Pindo Mirador biological station, Ecuador
Efeito da fase de sucessão florestal sobre a relação orquídea-epífita na estação biológica de Pindo Mirador, Equador
Christopher Oswaldo Paredes Ulloa1* ![]() https://orcid.org/0000-0002-2087-5694
https://orcid.org/0000-0002-2087-5694
Jorge Ferro-Díaz2 ![]() https://orcid.org/0000-0001-8101-7442
https://orcid.org/0000-0001-8101-7442
Pablo Lozano Carpio3 ![]() https://orcid.org/0000-0003-0857-8141
https://orcid.org/0000-0003-0857-8141
1Gobierno Autónomo Descentralizado Provincial de Pastaza (GADPPZ). Ecuador.
2Centro de Investigaciones y Servicios Ambientales ECOVIDA, Departamento Investigaciones, Ministerio de Ciencia, Tecnología y Medio Ambiente. Pinar del Río, Cuba.
3Universidad Estatal Amazónica, Centro de Investigación y Postgrado para la Conservación Amazónica CIPCA, Herbario ECUAMZ, Puyo, Pastaza. Ecuador.
*Autor para la correspondencia: chris9enginer@gmail.com
Recibido:08/11/2020.
Aprobado:12/01/2021.
RESUMEN
La diversidad de epífitas en los ecosistemas forestales está estrechamente relacionada con la estructura del bosque; sin embargo, en la Estación Biológica Pindo Mirador, en Ecuador, no existen estudios que documenten esa interacción. Se establecieron ocho parcelas de 0,1 ha cada una, estando cuatro en el bosque secundario y otras cuatro en el primario con el objetivo de evaluar el efecto del estadio sucesional actual del bosque sobre la relación hospederos - orquídeas epífitas en dicha estación, a partir del comportamiento de factores de la estructura forestal y la diversidad de orquídeas. Todas las orquídeas epífitas presentes y los árboles hospederos fueron registradas a partir de la medición de sus diámetros y alturas (total y del fuste), de comparaciones de la riqueza y abundancia por estadio y por hospederos, además de aplicarse un Análisis de Correspondencia Canónico entre variables de la estructura del bosque y los registros de epífitas por parcelas. Se detectó que el principal rasgo de la estructura horizontal del epifitismo orquideológico es la mayor abundancia y menor riqueza en el bosque secundario, ocurriendo lo contrario en el primario. Con respecto a la estructura vertical, una mayor abundancia de orquídeas en la parte más alta del fuste de los hospederos en el bosque secundario, mientras que en el primario predominan en la base de las ramas principales, concluyéndose que el estadio sucesional del bosque afecta tanto a la riqueza como a la abundancia de orquídeas sobre la base de los cambios que experimenta la estructura forestal, en cada uno de ellos.
Palabras clave: Altura de fuste; Apertura del dosel; Bosque primario; Bosque secundario; Clases diamétricas.
ABSTRACT
Epiphyte diversity in forest ecosystems is closely related to forest structure; however, in the Pindo Mirador Biological Station in Ecuador, there are no studies documenting this interaction. Eight plots of 0.1 ha each were established, four in the secondary forest and four in the primary forest, with the objective of evaluating the effect of the current successional stage of the forest on the host-epiphyte orchid relationship in that station, based on the behavior of forest structure factors and orchid diversity. All epiphytic orchids present and host trees were recorded by measuring their diameters and heights (total and stem), comparing richness and abundance by stage and host, and applying a Canonical Correspondence Analysis between forest structure variables and epiphyte records by plots. It was detected that the main feature of the horizontal structure of orchid epiphytism is the higher abundance and lower richness in the secondary forest, occurring the opposite in the primary forest. With respect to the vertical structure, a greater abundance of orchids in the highest part of the stem of the hosts in the secondary forest, while in the primary forest they predominate at the base of the main branches, concluding that the successional stage of the forest affects both the richness and abundance of orchids on the basis of the changes experienced by the forest structure, in each of them.
Keywords: Stem height; Canopy opening; Primary forest; Secondary forest; Diameter classes.
RESUMO
A diversidade epífita nos ecossistemas florestais está intimamente relacionada com a estrutura florestal; contudo, na Estação Biológica de Pindo Mirador, Equador, não existem estudos que documentem esta interação. Foram estabelecidas oito parcelas de 0,1 ha cada, quatro em floresta secundária e quatro em floresta primária, a fim de avaliar o efeito da sucessão atual da floresta sobre a relação orquídea-hóspede naquela estação, com base no comportamento dos fatores de estrutura florestal e diversidade de orquídeas. Todas as orquídeas epífitas presentes e as árvores hospedeiras foram registadas a partir da medição dos seus diâmetros e alturas (total e caule), comparações de riqueza e abundância por fase e por hospedeiro, e foi aplicada uma Análise de Correspondência Canónica entre as variáveis da estrutura florestal e os registos epífitos por parcela. Foi constatado que a principal característica da estrutura horizontal do epífito das orquídeas é a maior abundância e menor riqueza na floresta secundária, ocorrendo o oposto na floresta primária. No que respeita à estrutura vertical, uma maior abundância de orquídeas na parte mais alta do caule dos hospedeiros na floresta secundária, enquanto na floresta primária predominam na base dos ramos principais, concluindo que a fase sucessional da floresta afecta tanto a riqueza como a abundância de orquídeas com base nas mudanças experimentadas pela estrutura da floresta, em cada uma delas.
Palavras chave: Altura do eixo; Abertura do dossel; Floresta primária; Floresta secundária; Classes de diâmetros.
INTRODUCCIÓN
Los bosques tropicales están entre los ecosistemas de mayor biodiversidad del planeta, ya que albergan varias formas de vida. Esta alta diversidad es el resultado de los tipos de suelo, clima y condiciones biogeográficas en que se encuentran (Mudappa & Raman, 2007). Los bosques tropicales están sufriendo alteraciones estructurales recurrentes por los cambios ambientales globales (Stephenson et al., 2011).
El epifitismo constituye uno de los componentes que más destacan en los bosques tropicales, pues propicia el enriquecimiento de su biodiversidad, al posibilitar la ocupación de los diferentes estratos del bosque y con ello amplitud de ambientes para la mantención de la vida que no dependen directamente del suelo (Benzing, 1995; Leitman et al., 2015). Esto hace de los bosques tropicales uno de los ecosistemas más complejos de la biosfera (Besi et al., 2019).
Entender qué factores ambientales afectan las respuestas funcionales de las epífitas, ayudaría a establecer estrategias de manejo y conservación adecuadas para estos ecosistemas forestales, y en particular para su componente epifítico (Ferro y Delgado, 2013). Sin embargo, existen pocos estudios que posibiliten las actividades para el manejo adecuado de las epífitas en sus respectivos ecosistemas (Ferro, 2004; Hietz et al., 2006). En estos es recurrente el hecho de que se conoce poco sobre el impacto de deforestación sobre la diversidad de epífitas en bosques, y mucho menos sobre la ecología de epífitas de los bosques secundarios (Barthlott et al., 2001). Esto es importante porque las orquídeas son indicadoras ecológicas del incremento de temperaturas y aridez asociadas con la deforestación (Orta-Pozo, 2015).
La familia de plantas de epífitas del Ecuador con mayor diversidad documentada es Orchidaceae (más de 4 300 especies en el año 2014), es decir que casi una de cada cuatro especies de plantas que crecen en los hábitats silvestres del país son orquídeas, y representan más del 18 % del total de especies de orquídeas del mundo (García et al., 2014).
Esta familia aporta el mayor número de especies al fitoendemismo del país (1 707 especies), aproximadamente un tercio de las plantas endémicas son orquídeas (Endara et al., 2017). Sin embargo, no son precisas las cifras respecto a la cantidad de orquídeas epífitas y su distribución por ecosistemas, debido a que es insuficiente la difusión de reportes que abordan este aspecto, las que ocupan mayormente los ecosistemas forestales (Bravo, 2014).
En tal escenario del estado del conocimiento de la diversidad de orquídeas epífitas del Ecuador, cobra importancia estudios que no solo actualicen la información sobre dicha diversidad, sino que aporten nuevos argumentos sobre las interacciones con los hospederos, es por ello que los estudios en la Estación Biológica Pindo Mirador en la provincia de Pastaza, donde apenas existen investigaciones de las epifitas, son de gran relevancia para reforzar las acciones de protección en el área. En esta área, solo se ha reportado una lista de ocho especies de orquídeas recolectadas en su ecosistema forestal para el fomento de un orquideario de especies nativas en la propia Estación (Luzuriaga et al., 2017).
Dado el escaso conocimiento que se tiene de la diversidad de orquídeas epífitas del bosque de la Estación Biológica Pindo Mirador, y principalmente, sobre las características que tipifican la ecología de éstas, tanto en las áreas con bosque sin alteraciones antrópicas, como en la dinámica recuperativa que va experimentando el bosque en áreas con bosque secundario recuperado de impactos de talas intensas. Esta investigación tiene como objetivo: evaluar el efecto del estado sucesional del bosque sobre la relación hospederos-orquídeas epífitas a partir del comportamiento de factores de la estructura forestal y la distribución de la riqueza y abundancia de orquídeas epífitas en el bosque siempreverde piemontano en dos estadios sucesionales de la Estación biológica Pindo Mirador, Ecuador.
MATERIALES Y MÉTODOS
Área de estudio. La Estación Biológica Pindo Mirador (EBPM) tiene un área de 288,49 ha, ubicada en el cantón Mera de la provincia Pastaza, Ecuador, entre 1100 1300 m s.n.m., que pertenece a la cuenca del río Pastaza, y la microcuenca de los ríos Pindo Grande y su afluente el río Plata (Sucoshañay, 2016). El clima es mesotérmico, perhúmedo (Thornthwaite 1949) con precipitaciones medias anuales de 4 500 mm, temperaturas medias de 20ºC - 25ºC y humedad relativa promedio del 88 % (Luzuriaga, 2014).
La estación está ubicada en el bosque siempreverde piemontano del Norte de la Cordillera Oriental de los Andes, en el que se reconocen dos zonas principales, la Zona de Protección Permanente que se corresponde con el bosque primario, y la de Bosque Nativo con el bosque secundario, según Luzuriaga (2014) (Figura 1).
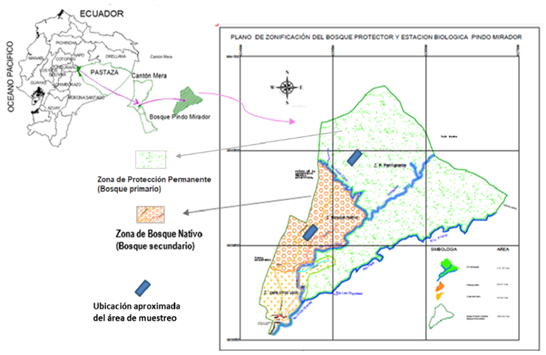
Figura 1. - Ubicación de las áreas de muestreo
Diseño del muestreo
Se establecieron ocho parcelas de 0,1 ha cada una, cubriendo un área de muestreo de 0,8 ha, ubicando cuatro en el bosque primario y cuatro en el secundario, las que se distribuyeron en cada estadio sucesional. Se utilizó para referirse a las parcelas en los análisis la nomenclatura siguiente: BP1, BP2, BP3 y BP4 son las cuatro parcelas del bosque primario, así como BS1, BS2, BS3 y BS4 representan las cuatro del bosque secundario.
Inventario de hospederos y orquídeas
Fueron inventariados todos los árboles hospederos de orquídeas con diámetros ≥ 10 cm de D1.30 según la metodología de Araújo et al., (2010). A estos se les determinó su diámetro del fuste, utilizando una cinta diamétrica y la altura total con un clinómetro Suunto, o por acceso hasta la copa con una cinta métrica de 50 m y, adicionalmente se midió la altura del fuste hasta el nacimiento de las primeras ramas. Se determinó el tipo de corteza de cada individuo arbóreo asumiendo dos tipos: corteza lisa y corteza rugosa (Migenis y Ackerman 1993).
Mediante exploración visual directa o a través de binoculares en cada árbol, se identificaron las orquídeas epífitas (holoepífitas según Kress, 1986), las que fueron determinadas a la observación, tanto en el fuste de los hospederos, como en sus ramas; los ajustes nomenclaturales se hicieron acorde a The Plant List (https://www.theplantlist.org/). La precisión de endemismo y categorías de amenazas reconocidas se obtuvo de Endara et al., (2017).
Se registraron las abundancias de orquídeas en cada hospedero por zonas de vida de éstos (forofíticas), acorde a su definición por Johansson (1974), modificadas para las dos primeras del fuste de los árboles, dada la variabilidad de altura observada, como fue asumida por Ferro (2004); estas fueron: Zona 1 (Z1): desde la base del fuste hasta la mitad del fuste, siempre por encima de los 1.30 m del suelo; Zona 2 (Z2): el resto del fuste desde la sección media de su altura hasta el nacimiento de las primeras ramas; Zona 3 (Z3): el primer tercio (basal) del largo de las ramas principales; Zona 4 (Z4): el tercio medio del largo de las ramas y Zona 5 (Z5): el último tercio o exterior del largo de las ramas.
Con el interés de evaluar posibles relaciones entre dimensiones de hospederos y la presencia y distribución de orquídeas epífitas, los registros de diámetros de los árboles fueron agrupados en cuatro clases: clase 1 (CD1): 10,0 a 19,9 cm, clase 2 (CD2): 20,0 a 29,9 cm, clase 3 (CD3): 30,0 a 39,9 cm y clase 4 (CD4): ≥ 40,0 cm; con el mismo fin se agruparon en cuatro rangos las alturas de los árboles, a saber: rango 1 (RH1): 2,00 a 7,99 m; rango 2 (RH2): 8,00 a 13,99 m; rango 3 (RH3): 14,00 a 19,99 m y rango 4 (RH4): 20,00 a 25,99 m; estas alturas se promediaron por parcelas para establecer una media del estrato arbóreo en cada unidad de muestreo.
Determinación de la cobertura del dosel
Se tomaron fotografías hemisféricas con lente ojo de pez, utilizando una cámara Cannon EOS 700D, siguiendo el protocolo propuesto por Kalácska et al. (2005), con ajustes para la fase intermedia de la sucesión que describen, reduciendo a 14 el número de fotografías por parcela, dada la diferencia en el tipo de ecosistema. Con el empleo del software GLA (Gap Light Analyzer) vers. 2.0 (Frazer et al., 1999) se procesaron las fotografías hemisféricas para obtener el porcentaje de apertura del dosel en cada parcela e incorporar los datos de esta variable a los análisis sobre sus posibles efectos en la distribución de orquídeas epífitas.
Análisis de los datos
Con el empleo del programa PAST vers. 3.01 (Hammer et al., 2001) se obtuvieron todos los estadísticos descriptivos y sus correspondientes representaciones gráficas, realizándose una comparación de medias de la riqueza, abundancia y cantidad de hospederos entre los dos estadios sucesionales, mediante una prueba de Kruskal-Wallis para p<0,05, misma prueba que se empleó para comparar los porcentajes de apertura del dosel entre los dos estadios sucesionales definidos; también se calculó la diversidad por parcelas según el índice de Simpson (1-D), obteniendo un criterio comparativo por estadio sucesional.
Se realizó un Análisis Canónico de Correspondencia (ACC) como sugiere Hill y Gauch (1980) para correlacionar una matriz de 10 variables ambientales (predictoras) con la abundancia por parcelas, de las especies que integran la comunidad de orquídeas epífitas, y mediante ello determinar cómo se ajustan las orquídeas en función de su relación con los efectos producidos por un conjunto de variables del ecosistema forestal; para ello se utilizó el software MVSP vers. 3.22demo- (Kovach Computing Services, 1985-2013: https://www.kovcomp.co.uk/downl2.html); las variables ambientales fueron: cantidad de los árboles hospederos por cada una de las cuatro clases diamétricas establecidas (criterio de preferencia o no por el grosor de hospederos), la abundancia de fustes en el bosque (criterio de influencia de la densidad de los árboles), promedio de apertura del dosel por parcelas (criterio de preferencia o no por ambientes más o menos iluminados), altura promedio de los arbóreos del bosque (criterio del efecto de la altura de la estratificación de la formación vegetal), altura promedio de fustes (criterio de efectos del soporte de los fustes), y los dos tipos de corteza (lisa y rugosa) de los fustes de hospederos (criterio de preferencia para fijación de epífitas).
RESULTADOS Y DISCUSIÓN
Riqueza y abundancia de orquídeas epífitas; efecto del estadio sucesional
Se identificaron 49 especies de orquídeas epífitas (ambos estadios sucesionales), 35 especies en el bosque secundario (BS), 48 en el bosque primario (BP); 14 especies que se localizaron en el BP no fueron observadas en el muestreo del BS, así como una sola especie no determinada del género Epidendrum presente únicamente en el BS. Tal riqueza pertenece a 20 géneros, siendo Maxillariella (10 especies) seguido de Elleanthus (8 especies) y Epidendrum (6 especies) los mejores representados. Del total, seis especies se reconocen como endémicas del Ecuador: Dichaea sodiroi Schltr, Epidendrum puyoense Hágsater & Dodson, Pleurothallis alveolata Luer, Sievekingia hirtzii Waldvogel, Sievekingia marsupialis Dodson y Stelis anolis Luer, las que a su vez poseen categoría de Amenazada: 4 Casi Amenazadas NT- y 2 Vulnerables V- según Endara et al., (2017).
La abundancia total de epífitas es de 2158 individuos, encontrándose una cifra más alta en el BS, en el que se contaron 1248, mientras que en el BP la cifra fue de 910. La diferenciación de riqueza de especies y abundancia por estadio sucesional se observa en la Figura 2 (A y B), donde, además, se reflejó el mayor rango de variación de los registros del BS tanto para la riqueza de especies, como para la abundancia de estas.
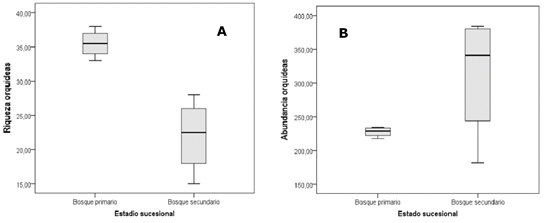
Figura 2. - Variación de la riqueza (A) y abundancia (B) de orquídeas epífitas por estadio sucesional
Los resultados encontrados para el bosque secundario son semejantes a los descritos por Eizmann y Zotz (2016) en parches de bosque secundario en Panamá, los que en su interpretación coinciden con el efecto de expansión rápida de epífitas en estado temprano de su dinámica post disturbio.
Se aprecia una mayor abundancia de orquídeas en el BS, donde también son más abundantes los hospederos; en el caso del BP su rasgo más importante es la mayor riqueza de especies de orquídeas epífitas. Un rasgo similar había sido encontrado por Barttlot et al., (2001) y Ferro (2004), los que exponen que, en una dinámica recuperativa, cuando el ecosistema forestal se encuentra en un estado más estable y maduro, la riqueza de las epífitas es mayor que en los estadios más próximos al disturbio, así como sus abundancias se regulan con el tránsito sucesional, algo que se ratifica con este estudio.
Distribución de árboles hospederos y riqueza y abundancia de orquídeas epífitas por estadios sucesionales
La Tabla 1 refleja el comportamiento del muestreo en cada parcela, y además se verifica que estas diferencias se mantienen en relación a la cantidad de árboles que hospedan tal reparto de la riqueza y abundancia.
Tabla 1. - Riqueza de orquídeas epífitas (S), su abundancia y cantidad de árboles que hospedan esas epífitas por estadio sucesional
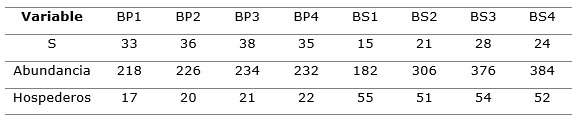
Los registros reportados de riqueza y abundancia de orquídeas epífitas en todas las parcelas del muestreo (Tabla 1), ofrece una clara diferenciación de cada una de las variables entre BP y BS, verificándose que al BP le caracteriza una mayor riqueza de especies, la que en promedio resultó en 35,5 especies para el BP y 22 para el BS, diferencia que es estadísticamente significativa (p=0,021); también se observa la variación respecto a la abundancia de orquídeas entre los dos estadios sucesionales, siendo mayor la del BS que la BP, diferencia numérica que no resultó estadísticamente significativa (p=0,248).
De estos resultados se puede interpretar que hay efecto del estadio sucesional para la riqueza de especies y para sus abundancias, siendo este efecto más marcado sobre la variación de la riqueza, la que se incrementa en la medida que el bosque es más maduro, sin embargo cuando está recuperándose de impactos relativamente recientes de la deforestación, la comunidad de orquídeas va estableciéndose, probablemente a partir del flujo proveniente de la matriz circundante de bosque primario, pero con un reparto más agresivo de sus abundancias, derivado de las condiciones que en este estadio va alcanzado en bosque en su trama de hospederos potenciales, afirmación que se sustenta en el hecho observado en la propia Tabla 1, en que en el BS una menor riqueza de especies ocupa mayor cantidad de árboles para sostenerse, razón que influye en el incremento de su abundancia respecto a la del BP.
La riqueza de especies concuerda con lo reportado por autores como Barthlott et al., (2001), que encontraron en bosques secundarios de los Andes venezolanos 36 especies de orquídeas epífitas en un área de 0,3 ha. Por su parte, Einzmann y Zotz (2016) hallaron 24 especies en siete parches de bosque secundario de Panamá; también se ajustan a los reportes de Hietz et al., (2006) y Besi et al., (2019). Guzmán-Jacob et al., (2020) demuestran que son importantes en la abundancia de epífitas las fuentes de bosques maduros próximos, todo lo cual se ajusta a la situación observada en Pindo Mirador; también se poyan los criterios expuestos en resultados como los de Mongosongo y Griffiths (2019), quienes argumentan que tanto la riqueza como abundancia de epífitas se incrementa en bosques más maduros debido a su mayor capacidad hospedante de los forófitos, lo cual hace que se preserven las variaciones requeridas para mayor cantidad de especies.
Dado el comportamiento descrito de la riqueza y abundancia en cada muestra para ambos estadios sucesionales, se verifica una mayor diversidad en el bosque primario con respecto al secundario (Figura 3), acorde a los valores del recíproco del índice de Simpson calculado (1/D), el que presenta valores siempre superiores a 0,92 en BP confirmando mayor probabilidad en la ocurrencia de individuos de una misma especie por el hecho de que la cantidad de estas es superior en la comunidad, algo que es inferior en el bosque secundario (1/D ≤ 0,87) , aun con sus más elevadas abundancias, debido a la diferencia significativa de su riqueza de especies respecto al bosque primario.
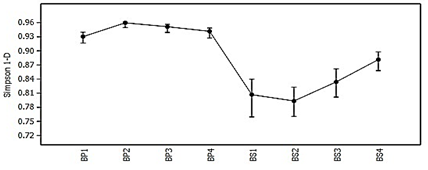
Figura 3. - Comportamiento de la diversidad según el recíproco del Índice de Simpson (1/D) cada estadio sucesional
Relación árboles-orquídeas por estadio sucesional
Con respecto a la riqueza arbórea y su contribución al epifitismo, como se aprecia en la Tabla 2, se observó que está tipificada por una mayor frecuencia de orquídeas epífitas en siete hospederos (seis especies más los árboles secos), de los cuales están en el bosque primario Alchornea latifolia Swart, Hyeronima oblonga (Tul.) Müll. Arg., Miconia splendens (Sw.) Griseb, Inga tessmanni Harms, Virola sp., Turpinia occidentalis (Sw.) G. Don y Viburnum ayavacense Kunth, con 52 que contienen orquídeas, de los 117 individuos observados; en tal diversidad de hospederos se encontraron 391 individuos de orquídeas epífitas y una riqueza promedio de 14 especies de orquídeas por árbol.
En la Tabla 2, se observa que en el caso del bosque secundario los hospederos con mayor número de especies de orquídeas fueron Miconia splendens (Sw.) Griseb, Piptocoma discolor (Kunth) Pruski, Palicourea guianensis Aubl, Hyeronima oblonga (Tul.) Müll. Arg., Bellucia pentamera Naudin., más los árboles secos, los que en total agruparon 267 individuos, y 174 de ellos fueron hospederos. En estos árboles hospederos del bosque secundario, se observó una abundancia de 823 orquídeas epífitas y una riqueza promedio de 16 especies por árbol.
Tabla 2. -Árboles con mayor contribución a la riqueza y abundancia de orquídeas epífitas en los dos estadios sucesionales evaluados
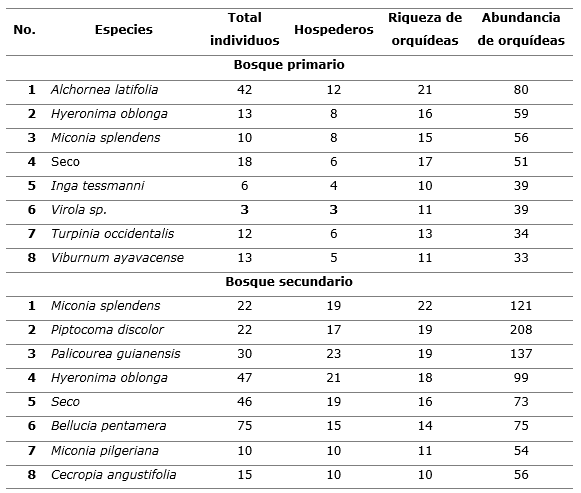
Distribución de orquídeas epífitas por clases diamétricas y zonas forofíticas; efecto del estadio sucesional
La distribución de orquídeas epífitas en relación a las clases diamétricas de sus hospederos (Figura 4 A y B) muestra la variación que se establece en cada estadio sucesional, observándose en el BP una mayor presencia en los árboles con diámetros de fustes entre 20 y 40 cm, característica que se ajusta a la dominancia en este estadio sucesional de árboles de superiores dimensiones a los del BS, como expone Luzuriaga et al., (2011), ya que en este último estadio predominan los que se encuentran entre 10 y 30 cm de D1,30, lo cual coincide con estudios en que se documentan incrementos de las dimensiones diamétricas con el avance de la dinámica sucesional, y reflejan que los estadios maduros ofrecen mayores capacidades en la arquitectura de hospederos para albergar al epifitísmo, lo cual es expuesto por Adhikari et al., (2017) y Hernández-Pérez et al., (2018).
Por su parte, en la distribución vertical de las orquídeas epífitas acorde a su ubicación en cada una de las cinco zonas forofíticas (Figura 4 C y D) se confirma el efecto del estadio sucesional, el que influye en la posición dominante en la sección superior de los fustes (en bosque secundario), o en la base de las ramas principales del dosel (en el bosque primario).
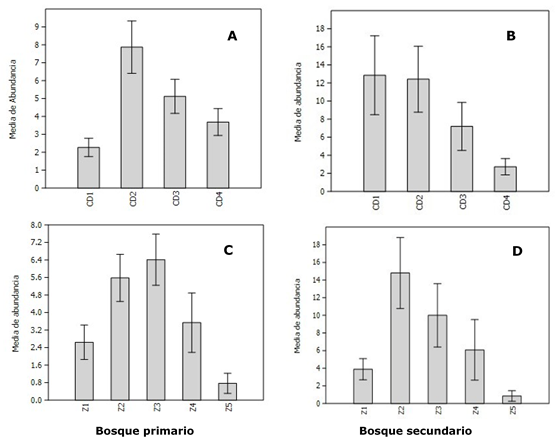
Figura 4. - Distribución de orquídeas epífitas según clases diamétricas (A y B) y zonas
forofíticas (C y D)
en los dos estadios sucesionales evaluados (bosque primario y bosque secundario)
Apertura del dosel
El análisis del porcentaje de apertura del dosel expone que en el BP el dosel es más cerrado que en el BS (Figura 5), lo que es estadísticamente significativo (p=0,0209); ello sugiere que el efecto del estadio sucesional sobre la distribución vertical del epifitismo orquideológico en Pindo Mirador está relacionado con la estructura del dosel del bosque, provocando que las orquídeas se concentren más en una zona de protección del dosel como la Z3, sin embargo, al ser este más abierto en el BS, las epífitas descienden, aprovechando la mayor protección de la penetración de radiación solar. Tal perspectiva sustenta un patrón semejante obtenido por Hernández-Pérez et al., (2018) y expuesto por Johansson (1974) acerca del efecto de las condiciones ambientales bajo el dosel acorde al tiempo en que va cambiando la estructura del bosque en su dinámica sucesional.
Figura 5. - Distribución del porcentaje de apertura del dosel en los dos estadios
sucesionales
(Bosque primario-Bprim- y Bosque secundario-Bsec-)
Correspondencia entre variables de la estructura del bosque y las parcelas, según la riqueza y abundancia de orquídeas epífitas observadas
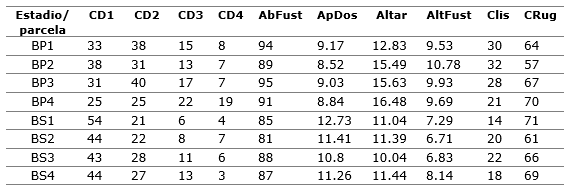
Las variables estructurales del bosque (predictoras) asumidas para ser correlacionadas con el muestreo de orquídeas epífitas (sitios de muestreo) en cada estadio sucesional (Tabla 3) representan un medidor de cómo los cambios de la estructura del bosque en ambos estadios sucesionales se relaciona con la variación del epifitismo orquideológico, pues algunas de ellas exponen cambios de uno a otro estadio (Tabla 3), entre las que se reconocen como más claras a las alturas de árboles, incluyendo sus fustes, el por ciento de apertura del dosel y los dos tipos de corteza de los árboles asumidos.
Tabla 3. - Valores de las variables de la estructura del bosque asumidas en el ACC por parcelas y por estadio sucesional, donde CD1 es la Clase Diamétrica 1, CD2 es la Clase Diamétrica 2, CD3 es la Clase Diamétrica 3, CD4 es la Clase Diamétrica 4, AbFust en la abundancia de fustes, ApDos es la apertura del dosel en por ciento, AltArb es la altura de árboles (promedio), AltFust es altura de fustes (promedio), Clis es corteza lisa y Crug es corteza rugosa
En el análisis de correspondencia canónico (ACC), se aprecia que los dos primeros ejes muestran una solución relativamente buena a la ordenación de especies y parcelas en función de las 10 variables ambientales, los que explican el 58,78 % de la variabilidad total (Tabla 4), la que incluido un tercer eje representa el acumulado de 72,09 %
Tabla 4. - Resultados del análisis canónico de correspondencia (ACC) de las ocho parcelas por estadios sucesional en función de las variables ambientales

En el gráfico biplot, se representa la proyección de variables ambientales, y parcelas de muestreo por estadio sucesional, se aprecian dos grupos, uno relacionado con el bosque secundario, donde se observa la dispersión del muestreo, poco efecto de variables; influencia de aquellas más vinculadas a una dinámica próxima a los impactos de las talas forestales como el predominio de arbóreos de menor dimensión diamétrica (CD1) y un mayor porcentaje en la apertura del dosel (ApDos) (Figura 6).
Por su parte, en el bosque primario hay un mayor efecto de variables relacionadas con una estructura de bosque maduro (mayor altura de los árboles y sus fustes, la abundancia de los arbóreos y las mayores dimensiones diamétricas de sus fustes).
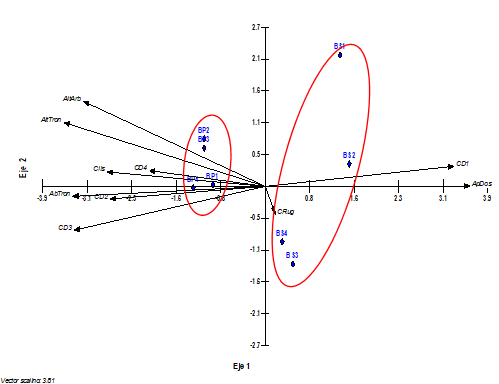
Figura 6. - Proyección de las variables ambientales y las parcelas del muestreo de orquídeas epífitas según el ACC. Las variables ambientales se muestran como flechas mientras que las parcelas como triángulos en color azul, donde BP y BS corresponde al estadio sucesional y el número acompañante al de las parcelas del muestreo
Los resultados del análisis anterior son semejantes con trabajos realizados por Krömer y Gradstein, (2003); Hietz et al., (2006); Flores-Palacios (2011) y García-Franco, 2008 y Larrea y Werner, (2010) los que coinciden que los cambios ambientales ocasionados por la eliminación o modificación de la estructura del bosque natural causan cambios en la composición, abundancia relativa, distribución espacial y respuestas funcionales de las epífitas.
Los resultados muestran el estadio sucesional representa una estructura que varía según persista o no la proximidad de impactos de talas intensas en el bosque. Benzing (1995) y Köster et al. (2009) fundamentaron que los cambios sucesionales en ecosistemas forestales afectan a las comunidades de epífitas de éstos, principalmente por variaciones en la estructura del dosel que influye en el régimen lumínico, la humedad y temperatura bajo éste; hecho que apoya la diferenciación observada en Pindo Mirador, y se confirma así un efecto del estadio sucesional sobre tales condiciones ambientales dentro del bosque, y en consecuencia en la caracterización de su epifitismo orquideológico.
CONCLUSIONES
El estado sucesional en que se encuentra el bosque afecta tanto a la riqueza como a la abundancia de orquídeas epífitas, con mayor relevancia en su distribución vertical, siendo más alta la contribución de las variaciones de altura de los fustes y ramificaciones principales de los árboles.
Las principales características estructurales de la comunidad de orquídeas se tipifican por ser más alta la abundancia y menor la riqueza de especies en el BS que en BP, ubicadas en sus hospederos de acuerdo a los árboles dominantes y mayormente, en la parte alta de los fustes en el BS, y en la base de las ramas principales en el BP.
REFERENCIA BIBLIOGRÁFICAS
ADHIKARI, Y.P., FISCHER, A., FISCHER, H.S., ROKAYA, M.B., BHATTARAI, P. y GRUPPE, A., 2017. Diversity, composition and host-species relationships of epiphytic orchids and ferns in two forests in Nepal. Journal of Mountain Science [en línea], vol. 14, no. 6, pp. 1065-1075. [Consulta: 14 de mayo de 2020]. ISSN 1993-0321. DOI 10.1007/s11629-016-4194-x. Disponible en: https://doi.org/10.1007/s11629-016-4194-x.
BARTHLOTT, W., SCHMIT-NEUERBURG, V., NIEDER, J. y ENGWALD, S., 2001. Diversity and abundance of vascular epiphytes: a comparison of secondary vegetation and primary montane rain forest in the Venezuelan Andes. Plant Ecology [en línea], vol. 152, no. 2, pp. 145-156. [Consulta: 14 de mayo de 2020]. ISSN 1573-5052. DOI 10.1023/A:1011483901452. Disponible en: https://doi.org/10.1023/A:1011483901452.
BENZING, D.H., 1995. THE PHYSICAL MOSAIC AND PLANT VARIETY IN FOREST CANOPIES. Selbyana [en línea], vol. 16, no. 2, pp. 159-168. [Consulta: 14 de mayo de 2020]. ISSN 0361-185X. Disponible en: https://www.jstor.org/stable/41759902.
BRAVO, E., 2014. La Biodiversidad en el Ecuador [en línea]. Ecuador: Editorial Universitaria Abya-Yala. [Consulta: 16 julio 2020]. Disponible en: http://190.57.147.202:90/jspui/bitstream /123456789/303/1/La%20Biodiversidad.pdf.
EINZMANN, H.J.R. y ZOTZ, G., 2016. How Diverse are Epiphyte Assemblages in Plantations and Secondary Forests in Tropical Lowlands? Tropical Conservation Science [en línea], vol. 9, no. 2, pp. 629-647. [Consulta: 17 julio 2020]. ISSN 1940-0829. DOI 10.1177/194008291600900205. Disponible en: https://doi.org/10.1177/194008291600900205.
ENDARA, L., HIRTZ, A. y JOST, L., 2017. Libro Rojo de Plantas Endémicas del Ecuador. Quito, Ecuador: Publicaciones del Herbario QCA, Pontificia Universidad Católica del Ecuador.
ENTALAI, E., NIKONG, D., MUSTAFA, M. y GO, R., 2019. Orchid diversity in antropogenic-induced degraded tropical rainforest, an extrapolation towards conservation. Lankesteriana [en línea], vol. 19, no. 2. DOI 10.15517/lank.v19i2.38775. Disponible en: https://www.researchgate.net/publication/335434623_Orchid_diversity_in_antropogenic-induced_degraded_tropical_rainforest_an_extrapolation_towards_conservation.
FERRO DÍAZ, J., 2004. Efecto del aprovechamiento forestal sobre la estructura y dinámica de la comunidad de epifitas vasculares del bosque semideciduo notófilo de la Península de Guanahacabibes. Tesis en opción al grado científico Doctor en Ciencias Forestales. Pinar del Río, Cuba: Universidad de Pinar del Río «Hermanos Saíz Montes de Oca».
FERRO, J. y DELGADO, F., 2013. Dinámica post disturbio de claros del dosel en el bosque tropical seco semideciduo de la Península de Guanahacabibes, Cuba; su relación con la abundancia de epífitas vasculares. En: L. FERNÁNDEZ y A. VOLPEDO (eds.), Evaluación de los cambios de estado de ecosistemas degradados de Iberoamérica [en línea]. Buenos Aires, Argentina: Programa Iberoamericano de Ciencia y Tecnología para el Desarrollo CYTED, pp. 200-213. Disponible en: https://es.scribd.com/document/327975559/Evaluacion-de-Los-Cambios-de-Estado-en-Ecosistemas-Degradados-de-Iberoamerica.
FLORES-PALACIOS, A., GARCÍA-FRANCO, J.G., VALENCIA-DÍAZ, S., SOLÍS-MONTERO, L. y CRUZ-ANGÓN, A., 2011. Diversidad y conservación de plantas epífitas vasculares en el centro del Estado. En: COMISIÓN NACIONAL PARA EL CONOCIMIENTO Y USO DE LA BIODIVERSIDAD (ed.), La Biodiversidad en Veracruz: Estudio de Estado [en línea]. México: Comisión Nacional para el Conocimiento y Uso de la Biodiversidad, Universidad Veracruzana : Instituto de Ecología, pp. 493-501. ISBN 978-607-7607-50-2. Disponible en: http://bibliotecasibe.ecosur.mx/sibe/book/000051158.
FRAZER, G.W., CANHAM, C.D. y LERTZMAN, K.P., 1999. Gap Light Analyzer (GLA): Imaging software to extract canopy structure and gap light transmission indices from true-colour fisheye photographs, user’s manual and program documentation. [en línea]. Millbrook, New York: University, Burnaby, British Columbia, and the Institute of Ecosystem Studies. Disponible en: http://rem-main.rem.sfu.ca/downloads/Forestry/GLAV2UsersManual.pdf.
GARCÍA, M., PARRA, D. y MENA, P., 2014. El país de la biodiversidad: Ecuador. Fundación Botánica de los Andes. Quito, Ecuador: Ministerio del Ambiente y Fundación EcoFondo.
GARCÍA-FRANCO, J.G. y TOLEDO, T., 2008. Epífitas vasculares: bromelias y orquídeas. En: R. MANSON, S. HERNÁNDEZ-ORTIZ y K. MEHLTRETER (eds.), Agroecosistemas cafetaleros de Veracruz. Biodiversidad manejo y conservación [en línea]. México: Instituto de Ecología/Instituto Nacional de Ecología, pp. 69-82. Disponible en: http://www2.inecc.gob.mx/publicaciones2/libros/542/cap5.pdf.
GUZMÁN‐JACOB, V., ZOTZ, G., CRAVEN, D., TAYLOR, A., KRÖMER, T., MONGE‐GONZÁLEZ, M.L. y KREFT, H., 2020. Effects of forest-use intensity on vascular epiphyte diversity along an elevational gradient. Diversity and Distributions [en línea], vol. 26, no. 1, pp. 4-15. [Consulta: 8 marzo 2021]. ISSN 1472-4642. DOI https://doi.org/10.1111/ddi.12992. Disponible en: https://onlinelibrary.wiley.com/doi/abs/10.1111/ddi.12992.
HAMMER, O., HARPER, D. y RYAN, P., 2001. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontologia Electronica [en línea], vol. 4, pp. 1-9. Disponible en: https://www.researchgate.net/publication/259640226_PAST_Paleontological_Statistics_Software_Package_for_Education_and_Data_Analysis.
HERNÁNDEZ-PÉREZ, E., SOLANO, E., RÍOS-GÓMEZ, R., HERNÁNDEZ-PÉREZ, E., SOLANO, E. y RÍOS-GÓMEZ, R., 2018. Host affinity and vertical distribution of epiphytic orchids in a montane cloud forest in southern Mexico. Botanical Sciences [en línea], vol. 96, no. 2, pp. 200-217. [Consulta: 6 de agosto de 2020]. ISSN 2007-4298. DOI 10.17129/botsci.1869. Disponible en: http://www.scielo.org.mx/scielo.php?script=sci_abstract&pid=S2007-42982018000200200&lng=es&nrm=iso&tlng=en.
HIETZ, P., BUCHBERGER, G. y WINKLER, M., 2006. Effect of forest disturbance on abundance and distribution of epiphytic bromeliads and orchids. Ecotropica [en línea], vol. 12, pp. 103-112. Disponible en: https://www.researchgate.net/publication/236685338_Effect_of_forest_disturbance_on_abundance_and_distribution_of_epiphytic_bromeliads_and_orchids.
HILL, M.O. y GAUCH, H.G., 1980. Detrended correspondence analysis: An improved ordination technique. Vegetatio [en línea], vol. 42, no. 1, pp. 47-58. [Consulta: 18 de junio de 2020]. ISSN 1573-5052. DOI 10.1007/BF00048870. Disponible en: https://doi.org/10.1007/BF00048870.
JOHANSSON, D., 1974. Ecology of vascular epiphytes in West African rain forest [en línea]. S.l.: Acta Phytogeographica Suecica. [Consulta: 16 julio 2020]. Disponible en: http://www.diva-portal.org/smash/get/diva2:565496/FULLTEXT01.pdf.
KALÁCSKA, M., CALVO-ALVARADO, J.C. y SÁNCHEZ-AZOFEIFA, G.A., 2005. Calibration and assessment of seasonal changes in leaf area index of a tropical dry forest in different stages of succession. Tree Physiology [en línea], vol. 25, no. 6, pp. 733-744. ISSN 0829-318X. DOI 10.1093/treephys/25.6.733. Disponible en: https://pubmed.ncbi.nlm.nih.gov/15805093/.
KÖSTER N, et al., 2009. Conservation of epiphyte diversity in an Andean landscape transformed by human land use. Conservation Biology : the Journal of the Society for Conservation Biology [en línea], vol. 23, no. 4, pp. 911-919. [Consulta: 18 de junio de 2020]. ISSN 0888-8892, 1523-1739. DOI 10.1111/j.1523-1739.2008.01164.x. Disponible en: https://europepmc.org/article/med/19210304.
KRÖMER, T. y GRADSTEIN, S.R., 2003. Species Richness of Vascular Epiphytes in Two Primary Forests and Fallows in the Bolivian Andes. Selbyana [en línea], vol. 24, no. 2, pp. 190-195. [Consulta: 18 de junio de 2020]. ISSN 0361-185X. Disponible en: https://www.jstor.org/stable/41760132.
LARREA, M.L. y WERNER, F.A., 2010. Response of vascular epiphyte diversity to different land-use intensities in a neotropical montane wet forest. Forest Ecology and Management [en línea], vol. 260, no. 11, pp. 1950-1955. [Consulta: 8 marzo 2021]. ISSN 0378-1127. DOI 10.1016/j.foreco.2010.08.029. Disponible en: https://www.sciencedirect.com/science/article/pii/S0378112710004883
LEITMAN, P., AMORIM, A.M., SANSEVERO, J.B.B. y FORZZA, R.C., 2015. Floristic patterns of epiphytes in the Brazilian Atlantic Forest, a biodiversity hotspot. Botanical Journal of the Linnean Society [en línea], vol. 179, no. 4, pp. 587-601. [Consulta: 18 de junio de 2020]. ISSN 1095-8339. DOI https://doi.org/10.1111/boj.12342. Disponible en: https://onlinelibrary.wiley.com/doi/abs/10.1111/boj.12342.
LUZURIAGA, C., CUASAPAZ, C. y QUICHIMBO, G., 2011. INVENTARIO FORESTAL EN LA ESTACIÓN PINDO MIRADOR. Tsafiqui - Revista Científica en Ciencias Sociales [en línea], no. 2, pp. 83-107. [Consulta: 18 de junio de 2020]. ISSN 2602-8069. DOI 10.29019/tsafiqui.v0i2.211. Disponible en: https://revistas.ute.edu.ec/index.php/tsafiqui/article/view/211.
LUZURIAGA, C.X., QUICHIMBO, G.I. y BLANCO, J., 2017. Viverización de orquídeas epífitas como estrategia de conservación de especies autóctonas en los bosques amazónicos de Pastaza (Ecuador). Memorias del 7mo. Congreso Forestal Español [en línea]. Ecuador: Sociedad Española de Ciencias Forestales, Disponible en: http://7cfe.congresoforestal.es/content/viverizacion-de-orquideas-epifitas-como-estrategia-de-conservacion-de-especies-autoctonas-en.
LUZURIAGA Q, C.X., 2014. INVENTARIO FLORÍSTICO DEL BOSQUE QUE RODEA A LA LAGUNA EN LA ESTACIÓN BIOLÓGICA PINDO MIRADOR PASTAZA-ECUADOR. Tsafiqui - Revista Científica en Ciencias Sociales [en línea], no. 6, pp. 15-25. [Consulta: 18 de junio de 2020]. ISSN 2602-8069. DOI 10.29019/tsafiqui.v0i6.228. Disponible en: https://revistas.ute.edu.ec/index.php/tsafiqui/article/view/228.
MIGENIS, L.E. y ACKERMAN, J.D., 1993. Orchid—phorophyte relationships in a forest watershed in Puerto Rico. Journal of Tropical Ecology [en línea], vol. 9, no. 2, pp. 231-240. [Consulta: 8 marzo 2021]. ISSN 1469-7831, 0266-4674. DOI 10.1017/S0266467400007227. Disponible en: https://www.cambridge.org/core/journals/journal-of-tropical-ecology/article/abs/orchidphorophyte-relationships-in-a-forest-watershed-in-puerto-rico/6B4579E8A64C56E734836F38ADEEDDC9#.
MUDAPPA, D., RAMAN, T.R.S., SHAHABUDDIN, G. y RANGARAJAN, M., 2007. Rainforest restoration and wildlife conservation on private lands in the Western Ghats. Making conservation work [en línea]. India: Permanent Black, Ranikhet, pp. 210-240. Disponible en: https://www.researchgate.net/publication/258699793_Rainforest_restoration_and_wildlife_conservation_on_private_lands_in_the_Western_Ghats.
STEPHENSON, N.L., MANTGEM, P.J. van, BUNN, A.G., BRUNER, H., HARMON, M.E., O’CONNELL, K.B., URBAN, D.L. y FRANKLIN, J.F., 2011. Causes and implications of the correlation between forest productivity and tree mortality rates. Ecological Monographs [en línea], vol. 81, no. 4, pp. 527-555. [Consulta: 18 de junio de 2020]. ISSN 1557-7015. DOI https://doi.org/10.1890/10-1077.1. Disponible en: https://esajournals.onlinelibrary.wiley.com/doi/abs/10.1890/10-1077.1.
SUCOSHAÑAY, D.J., 2016. Propuesta para el ordenamiento ambiental de la Cuenca del Río Puyo, en la amazonía ecuatoriana [en línea]. Tesis doctoral. La Habana, Cuba: Facultad de Geografía, Universidad de La Habana. Disponible en: http://eduniv.reduniv.edu.cu/index.php?page=13&id=272&db=1.
THORNTHWAITE, C.W., BIEL, E.R., CHURCH, P.E., JACOBS, W.C., LANDSBERG, H., LEIGHLY, J.B. y HAFSTAD, K., 1949. Report of the Committee on Climatology, 1947–1948. Eos, Transactions American Geophysical Union [en línea], vol. 30, no. 3, pp. 439-443. [Consulta: 18 de junio de 2020]. ISSN 2324-9250. DOI https://doi.org/10.1029/TR030i003p00439. Disponible en: https://agupubs.onlinelibrary.wiley.com/doi/abs/10.1029/TR030i003p00439.
Conflicto de intereses:
Los autores declaran no tener conflictos de intereses.
Contribución de los autores:
Christopher Oswaldo Paredes Ulloa: Concepción de la idea, búsqueda y revisión de literatura, confección de instrumentos, aplicación de instrumentos, recopilación de la información resultado de los instrumentos aplicados, análisis estadístico, confección de tablas, gráficos e imágenes, confección de base de datos, redacción del original (primera versión), revisión y versión final del artículo, coordinador de la autoría, traducción de términos o información obtenida, revisión de la aplicación de la norma bibliográfica aplicada.
Jorge Ferro-Díaz: Concepción de la idea, búsqueda y revisión de literatura, confección de instrumentos, aplicación de instrumentos, recopilación de la información resultado de los instrumentos aplicados, análisis estadístico, confección de tablas, gráficos e imágenes, confección de base de datos, asesoramiento general por la temática abordada, revisión y versión final del artículo, corrección del artículo, traducción de términos o información obtenida.
Pablo Lozano Carpio: Búsqueda y revisión de literatura, confección de instrumentos, confección de tablas, gráficos e imágenes, asesoramiento general por la temática abordada, revisión y versión final del artículo, corrección del artículo.
![]()
Esta obra está bajo una licencia de Creative Commons Reconocimiento-NoComercial
4.0 Internacional.
Copyright (c) 2021 Christopher Oswaldo Paredes Ulloa, Jorge Ferro-Díaz, Pablo Lozano Carpio