Translated from the original in spanish
Determination of the optimal photographic conditions for the anatomical characterization of ten timber species in Costa Rica
Determinación de las condiciones fotográficas óptimas para la caracterización anatómica de diez especies maderables de Costa Rica
Determinação das condições fotográficas ideais para a caracterização anatómica de dez espécies de madeira na Costa Rica
Juan Carlos Valverde1* ![]() https://orcid.org/0000-0002-3181-1346
https://orcid.org/0000-0002-3181-1346
Dagoberto Arias1 ![]() https://orcid.org/0000-0002-3056-9172
https://orcid.org/0000-0002-3056-9172
Erick Mata1 ![]() https://orcid.org/0000-0001-5471-164X
https://orcid.org/0000-0001-5471-164X
Geovanni Figueroa1 ![]() https://orcid.org/0000-0003-3788-3786
https://orcid.org/0000-0003-3788-3786
Nelson Zamora1 ![]() https://orcid.org/0000-0002-4358-6123
https://orcid.org/0000-0002-4358-6123
1Tecnológico de Costa Rica. Costa Rica.
*Correspondence author: jcvalverde@itcr.ac.cr
Received: June 14th, 2020.
Approved: July 13 th, 2020.
ABSTRACT
There is an absence of studies on tropical timber species that show the optimization in quantity and dimensions of wood photography in anatomical studies, an aspect that restricts the statistical representativeness in anatomical studies in tropical species. The study presented as an objective to determine the dimensions and minimum quantity of macroscopic photographs that can be represented in tropical species and to analyze the relationship between photographic dimensions and anatomical characteristics. Ten tree species were selected for this purpose; eight to fifteen cylindrical wood specimen of 12.5 mm diameter and 75 mm length were collected from each species, which were dried and photographed on the transverse side. The results showed a variation from 19 to 35 photographs as minimum quantity per species, being Spondias mombin the species with the highest requirement of photographs with 35, while Caryocar costarricense the one with the lowest amount of minimum photographs; furthermore, the optimal common dimension of photographs was 1.25 x 1.25 mm, being Peltogyne purpurea the species that could be worked with even lower dimensions. The combination of the minimum number of photographs and the optimal dimension allowed to be representative and reproducible in more than 90 %. Finally, a correlation was found between the size of the image and the diameter of the vessels; the ratio was that as the diameter of the vessels is smaller, it is possible to work with smaller photographs with a ratio greater than 70 %.
Keywords: Wood anatomy; Microscopy; Tropical species.
RESUMEN
Existe una ausencia de estudios en especies maderables tropicales que muestren la optimización en cantidad y dimensiones de fotografía de madera en estudios anatómicos, aspecto que restringe la representatividad estadística en estudios anatómicos en especies tropicales. El estudio presentó como objetivo determinar la dimensiones y cantidad mínima de fotografías macroscópicas representables en especies tropicales y analizar la relación entre dimensiones fotográficas y características anatómicas. Para ello se seleccionaron diez especies arbóreas; de cada especie se colectaron de ocho a quince muestras cilíndricas de madera de 12,5 mm de diámetro y longitud de 75 mm, las cuales se secaron y fotografiaron en la cara trasversal. Los resultados mostraron una variación de 19 a 35 fotografías como cantidad mínima por especie, siendo Spondias mombin la especie con mayor requerimiento de fotografías con 35, mientras Caryocar costarricense la de menor cantidad de fotografías mínimas; además, la dimensión óptima común de fotografías fue de 1,25 x 1,25 mm, siendo Peltogyne purpurea la especie que se pudo trabajar con dimensiones todavía más bajas. La combinación de la cantidad de fotografías mínimas y la dimensión óptima permitió tener representatividades y reproducibilidades superiores al 90 %. Finalmente, se halló una correlación entre la dimensión de la imagen y el diámetro de vasos; la relación consistía en que conforme el diámetro de vasos es menor, es posible trabajar con fotografías de menor dimensión con una relación superior al 70 %.
Palabras clave: anatomía de la madera; microscopia; especies tropicales.
RESUMO
Há uma ausência de estudos sobre espécies de madeiras tropicais que mostram a optimização em quantidade e dimensões da fotografia em estudos anatómicos de madeira, um aspecto que restringe a representatividade estatística em estudos anatómicos em espécies tropicais. O estudo apresentado como objectivo para determinar as dimensões e quantidade mínima de fotografias macroscópicas que podem ser representadas em espécies tropicais e para analisar a relação entre dimensões fotográficas e características anatómicas. Dez espécies de árvores foram selecionadas para este fim; oito a quinze amostras cilíndricas de madeira de 12,5 mm de diâmetro e 75 mm de comprimento foram recolhidas de cada espécie, que foram secas e fotografadas no lado transversal. Os resultados mostraram uma variação de 19 a 35 fotografias como quantidade mínima por espécie, sendo Spondias mombin a espécie com maior exigência de fotografias com 35, enquanto Caryocar costarricense a espécie com menor quantidade de fotografias mínimas; além disso, a dimensão óptima comum das fotografias foi de 1,25 x 1,25 mm, sendo Peltogyne purpúrea a espécie que poderia ser trabalhada com dimensões ainda mais baixas. A combinação do número mínimo de fotografias e a dimensão óptima permitiu ter representatividades e produtividades superiores a 90 %. Finalmente, foi encontrada uma correlação entre o tamanho da imagem e o diâmetro dos vasos; a razão foi que, como o diâmetro dos vasos é menor, é possível trabalhar com fotografias menores com uma razão superior a 70 %.
Palavras-chave: Anatomia da madeira; Microscopia; Espécies tropicais.
INTRODUCTION
The stem is a relevant organ for plants, allowing the transport and administration of water, salts and nutrients, being fundamental for the growth, support and survival of individuals (Biondi and Rossi, 2015). In the specific case of woody species, the stem tends to form the xylem and phloem tissues that perform the function of storage and transport of water and nutrients in the tree (Sevanto et al., 2008). The xylem is the inactive tissue of the trunk, it is closer to the medulla of the tree and its main function is storage (Ayala-Usma et al., 2019); commercially, the xylem is known as wood and it is implemented as a constructive material in real estate buildings, furniture, firewood, among other uses (Campelo et al., 2018).
The wood can vary its physical and mechanical properties according to its origin (characteristics of the site where the tree grew) (Paz Pérez-Olvera and Dávalos-Sotelo, 2008) and genetics (Biondi and Rossi, 2015); however, there are aspects that remain homogeneous within each tree species that are their anatomical properties, aspects such as the distribution and shape of vessels, parenchyma, fibers, among others, tend to follow the same patterns in the species regardless of the site, growth conditions and genetics (Beyer et al, 2014), an aspect that has allowed the use of wood as a tool for species identification and has generated the creation of wood collections as a mechanism for consultation and comparison of samples, commonly called xylotheque (Machado et al., 2019).
One of the limitations of xylotheque is their limited accessibility by digital means such as virtual collections (Havinga et al., 2020), which limits their consultation by the public and their implementation in research due to the restriction in the number of samples (Malik et al., 2019). In addition, another limitation in the consultation of xylotheque is to define the minimum size of the sample necessary to make anatomical comparisons (Carlsson and Isaksson, 2018); many times, the tree species to be compared are alive or are samples of historical buildings, which limits the extraction of traditional samples without generating significant damage to the plant or building of origin.
Thus, it is necessary to start developing tests to define the minimum dimensional sample that allows anatomical analysis. Dantas and Cunha (2020) mention that traditionally cubic samples of 10 mm face have been used, which are implemented with microscope at 40 and 100X magnification for anatomical analysis; on the other hand, Sánchez-Costa et al., (2015) highlight the use of cubic samples of 20 mm face for macroscopic analysis at 10X and microscopic at 100X; Arzac et al., (2018) mention that samples of different sizes can be made depending on the error of the sample size, as the sample is smaller the representation error will increase, therefore it is not recommended to use cubic samples smaller than 4 mm in length. Micco et al. (2008), on the other hand, mention that the size of the sample will depend on the type and diameter of the vessel of the species under study, as the vessel is larger, the sample size will increase because it will be necessary a greater area to represent the anatomy of the species.
Brown (2013) mentions that defining optimal dimensions in sample digitizing not only optimizes the sample that is extracted from the tree wood, generating the minimum impact on the individual, but also allows for improving the training and learning processes in automatic species identification systems through algorithms, since macroscopic photography generally tends to be sectioned into smaller photos to increase the amount of images in the training process (Glabasnia and Hofmann, 2006); but, if this segmentation is very high, the representativeness of a species can be lost and one can fall into the error of having photographs that cover an entire vessel or absent vessels, which will induce a greater bias in the identification of the species (Ma and Huang, 2018).
Stepanova et al., (2013) mention that, for the creation of virtual species collections, there must be a significant sample of the species, with correct images that allow real identification and a selection of images that are representative or iconic of the species. In the case of macroscopic anatomy, it is necessary to determine a quantity of images that will give a minimum representativeness of the species, the determination of the minimum dimensions of the image and that will allow the representativeness and reproducibility of the species; however, the availability of these studies has been reduced at a global level.
In the case of tropical regions, the development of anatomical studies of tree species is very limited due to the wide diversity and variability of species, which makes it necessary, first, to generate protocols that are related to the region and that allow for an adequate study of the species (Brownson et al., 2020). Therefore, this study has set as an objective: to determine the minimum photographic dimensions to have statistical representativeness for anatomical macroscopic analysis, to define the minimum amount of macroscopic photographs that can be represented by species and to analyze the relationship between photographic dimensions and anatomical characteristics.
MATERIALS AND METHODS
Species and study sites
The study used ten tree species from forests on the Pacific Coast of Costa Rica. Specifically, they were collected from four locations (Figure 1) that were located at the Horizontes Experimental Forest Station in Guanacaste (10°42'10'' N, 85°33'12'' W) at an altitude of 120 m, forests in the region of Miramar, Puntarenas (10°01'29'' N, 84°14'04'' W) at an altitude of 270 m; In both regions the forests were characterized as dry with an annual water regime of less than 2 000 mm, with temperatures ranging from 27 to 34 °C and a defined dry season from December to May. Samples were also collected in the Osa Peninsula, Puntarenas; specifically in the Mogos forest (8°45'00'' N, 83°22'59'' W) at an altitude of 40 m and San Juan (8°39'02'' N, 83°27'53'' W) at an altitude of 35 m, sites that were characterised by a very short dry season (from January to April), with annual rainfall of 4 000 to 4 300 mm and annual temperature varying from 27 to 33 °C with an annual relative humidity of 70 to 90 %.
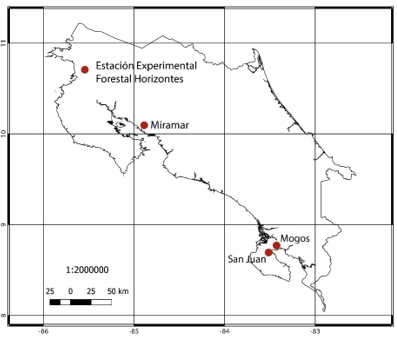
Figure 1. - Collection locations of the ten tree species in Costa Rica
From each species, no less than eight individuals were selected (Table 1), that have diameter at 1.3 m of 30 cm and with a distance between trees greater than 15 m (in order to increase the representativeness of the species on site).
Table 1. - Scientific names of the ten timber species
Process of material collection
For each individual in the study, a 75 mm long, 25 mm diameter sample of wood was obtained through non-destructive sampling using a spiral cork cutter type drill implemented in a 20 volt battery drill. The perforation was made in each tree at a height of 1.3 m above the ground, an area of 4 cm2 was cleaned, which was released from the bark and the drill bit was placed to drill the shaft, the drilling was done with a speed of 1 750 RPM; after the drilling was done with the full drill bit, the piece of wood was removed with a clamp and placed in a bag with a 15 % alcohol solution.
Material processing
Each cylindrical sample was segmented into five 13 mm long sub-cylinders, in which the cross faces were polished to make them as smooth as possible; each sub-cylinder was then manually converted into 12 mm face cubes with surfaces that were as smooth as possible and free of defects. They were then dried at a temperature of 65 °C for 72 hours so as not to have any drastic shrinkage of the wood to be exposed to high temperatures in a short time. After drying, the cubes were ground in order to have a cross-section as clean as possible, free of twists, drying defects or marks from the cutting process; the size of each face was 10 mm.
Photography procedure
The samples were photographed on their transverse side in a Celestron stereo-electronics scope, at 5 MP resolution and 20X magnification. The material was placed on a base bench and with a standard luminosity LED that has the integrated stereoscope, being used with an intensity of 150 lux, with a temperature of the light of 4 500 K; each sample was photographed four times, turning the sample 90° in each photography.
Anatomical analyses considered
Each photograph was processed with the "Image J" program and the density and average diameter of the vessels were estimated, as well as the vessel-fiber ratio. For this purpose, the Valverde and Arias (2018) methodology was implemented for tropical broadleaf tree species.
Statistical analysis
Firstly, a descriptive analysis of the anatomy of the wood samples on the cross-section of the ten species was carried out, analysing vessel density and vessel diameter. Subsequently, the optimal number of photographs per species was determined (implemented by reference the 10 x 10 mm image), for which a Cochran analysis of homogeneity of values was performed; until this analysis showed a variation greater than 0.05 it would show that the number of photographs would not be significant for the analysis. The optimal photographic dimension by species was then determined from both anatomical variables, using the minimum number of photographs and sectioning them into 10 x 10 mm, 5 x 5 mm, 2.5 x 2.5 mm, 1.25 x 1.25 mm, 0.62 x 0.62 mm, 0.31 x 0.31 mm, 0.17 x 0.17 mm and 0.08 x 0.08 mm images (Figure 2). The images were implemented to estimate the density and diameter of vessels and with Cochran's homogeneity of values test the point of variation was estimated.
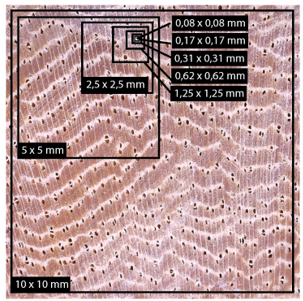
Figure 2. - Example of the photographic dimensions analysed for ten timber species
With regard to the determination of repeatability, the error of Cochran's test for homogeneity
of values was implemented; the formula was used:
(1- error)* 100 %. With regard to the reproducibility the formula was applied: ![]() where s is the standard deviation of the
analysed sample and V is the correction coefficient of Cochran's test performed. Finally, a Pearson
correlation analysis was performed with the optimum number and dimensions of the images to identify
the correspondence between the photographic dimension and the vessel diameter
where s is the standard deviation of the
analysed sample and V is the correction coefficient of Cochran's test performed. Finally, a Pearson
correlation analysis was performed with the optimum number and dimensions of the images to identify
the correspondence between the photographic dimension and the vessel diameter
All analyses were performed with a confidence level of 0.05 and the Origin Pro statistical program version 2020 was implemented.
RESULTS AND DISCUSSION
Initial anatomical characterization of the species
Table 2 presents the macroscopic characterization data of the ten-tree species under study. Vessel density ranged from 3.5 to 20.4 vessels mm-2, with E. cyclocarpum species presenting the lowest vessel density, while P. purpurea had the highest density with 20.4 vessels mm-2. Regarding the diameter of the vessels, it varied from 119 to 252 µm, being the species P. quinata the one with the lowest vessel diameter, while the species E. cyclocarpum showed the highest diameter.
The results shown are consistent with the studies developed by De La Paz Pérez-Olvera and Dávalos-Sotelo (2008) for six species that determined ranges of density and vessel diameter; only five species were found in that study and presented values with a similarity of 80 %, the variations obtained may be due to climatic, genetic and age variations of the individuals analyzed (Giachi et al., 2017), aspects that according to Kobayashi et al., (2019).
Table 2. - Macroscopic characterization of eight forest tree species in Costa Rica
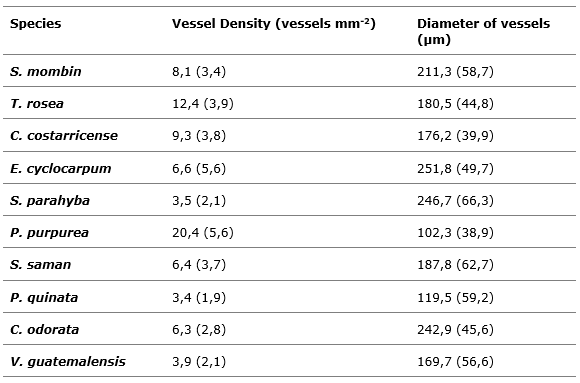
Note: values in brackets correspond to the standard deviation
Determination of the minimum number of images for species representativeness
In determining the number of minimum images per species, as a function of the variation in vessel density (Figure 3), the following characteristics were observed: as the number of images increased, the variation in vessel estimation decreased; the high values of vessel variation in a small number of images are due to the fact that many images only covered sections of the vessel, which made the heterogeneity of images wide. Variation in vessels in samples with few images meant that in most species the diameter of vessels tended to be overestimated, and this variation decreased as the number of images increased.
The study identified three groupings in terms of the number of photographs. The first grouping was formed by C. costarricense, T. rosea, P. purpurea, S. saman and P. quinata, which showed the least amount of images necessary to obtain a homogeneity of the error. The minimum number of images varied from 19 to 23 (Figure 3b, 3c, 3f, 3g and 3h); after that number of images there were no statistical differences with the increase in the number of photographs. The second grouping was made up of the species E. cyclocarpum, C. odorata and V. guatemalensis, which presented a minimum number of significant images of 25 to 30, after which the variation stabilized. Finally, the species S. mombin showed the highest number of images (35).
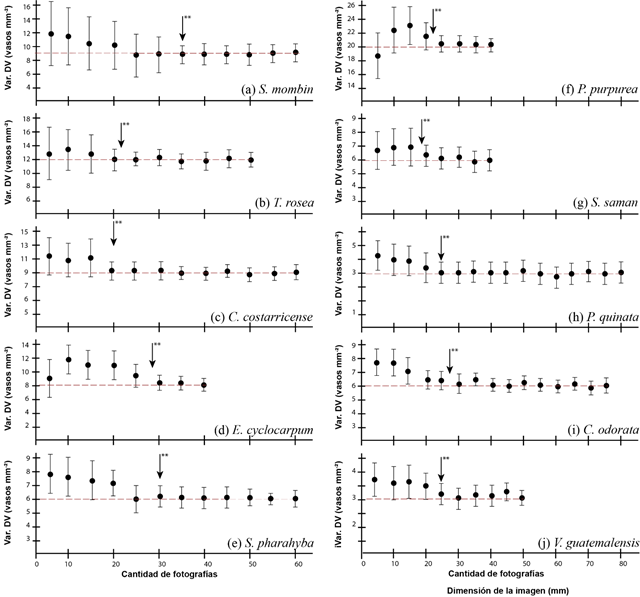
Figure 3. - Determination of the minimum significant amount of macroscopic wood images for ten tree species in Costa Rica
The poor functionality of the implementation of less than 19 images per species is due to the loss of representativeness of the species. Pacheco et al., (2020) mention that few representative images of a species generate an error in the anatomical data, since different parts of the anatomy of the species are found due to randomness effects and as the sample is increased there will be a greater representativeness of all the possible elements that make up the wood; thus, a greater set of information will be available. Studies carried out by Andrade Bueno et al., (2020) with Eucalyptus spp. species determined that anatomical analyses with less than five images are not very viable, due to the fact that errors can be made regarding the representativeness of the species; furthermore, only samples of juvenile wood can be taken, or only sapwood or heartwood sections, which affects the representation of the species.
Fioravanti et al., (2017) mention that once the quantity of optimal images per species has been determined, one should avoid working with lower quantities of images that can generate biases in statistical analyses or in representative descriptions of the species; aspects such as the distribution of the species, age, environmental conditions of development should be considered when taking the sample of images, since in many cases the use of a single origin or region can be inferred in the representation (Ayala-Usma et al, 2019), which is why not only the minimum number of samples should be considered, but also the distribution of individuals and their developmental conditions.
Determination of the minimum image size as a function of the vessel diameter
Regarding the determination of the optimal size of the image, depending on the diameter of the vessels (Figure 4), an optimal dimension of 1.25 x 1.25 mm was determined for most species, with T. rosea, C. costarricense and C. odorata having the optimal dimension of 0.62 x 0.62 mm, while with P. purpurea the dimension was 0.31 x 0.31 mm. In addition, it was determined that the dimensions below the optimal one overestimated the diameter of the vessels as a whole of a significant increase of the standard deviation, which influences an increase of the variation.
The determination of a minimum working dimension is fundamental to have a real representation of the image. Machado et al., (2019) point out that images with a very small dimension lose the representativeness of the anatomical elements; aspects such as vessels, parenchyma or fibers may have little representation in the image, generating bias when making anatomical calculations. Carlsson and Isaksson (2018) mention that in broadleaved species, care must be taken with the size of the image; small-sized images have a low capacity for species identification, due to the cutting of vessels and the low presence of anatomical elements within the image.
On the other hand, Brown et al., (2020) mention that the optimization of the size allows the development of optimized samples in the wood; thus, defining the size of a sample allows reducing the problem of making non-destructive samples or low impact samples to the study material.
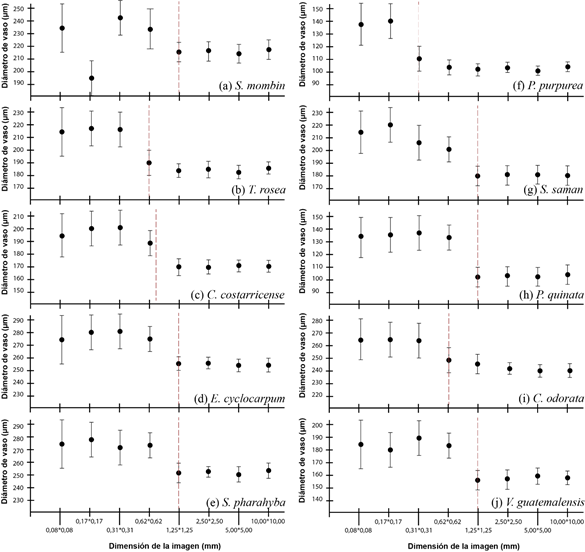
Figure 4. - Determination of the optimal size of macroscopic wood images as a function of vessel diameter for ten tree species in Costa Rica
Determination of minimum image size as a function of vessel density
With regard to the determination of the photographic dimension as a function of vessel density (Figure 5), according to the photograph, the density tended to be underestimated, the optimal size being 1.25 x 1.25 mm. For the species T. rosea, C. costerricensis and C. odorata the optimal size was 0.62 x 0.62 mm; on the other hand, for the species P. purpurea the optimal size was 0.31 x 0.31 mm. After the optimal dimension, the vessel density values had to stabilize and the error remained homogeneous.
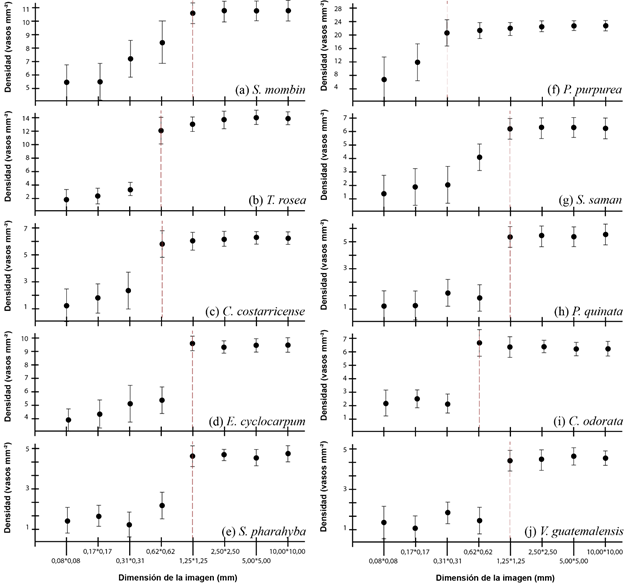
Figure 5. - Determination of the optimal size of macroscopic wood images as a function of vessel diameter for ten tree species in Costa Rica
Repeatability and reproducibility of optimal photo sampling conditions
When analyzing the repeatability and reproducibility of the optimal dimensions and number of images, values above 90 % were obtained for all species, which allowed to demonstrate the viability of the selected parameters; smaller quantities of photographs or lower photographic dimensions generated values below 60 %.
According to Sousa Fernandes et al., (2019) these aspects are relevant in studies of the representativeness of a species. As the ability to reproduce a study with the same conditions and similar results is given, real conditions will be available for the comparison of results. Copini et al., (2015) mention that in many cases it is complicated, due to deficiencies in methodologies, errors in data collection and manipulation procedures, omissions of steps that make the reproducibility of the experiment poor and the representativeness of the results possible.
The analyses showed that smaller dimensions than those determined made the variation in results wide, which increased the error and decreased the quality of the information, an aspect that Marcelo-Peña et al., (2019) consider deficient in studies with tree species, since biases in the anatomical values can be generated (Table 3).
Table 3. - Repeatability and reproducibility with the optimum quantity and dimensions
of the
photographs obtained for ten timber species in Costa Rica
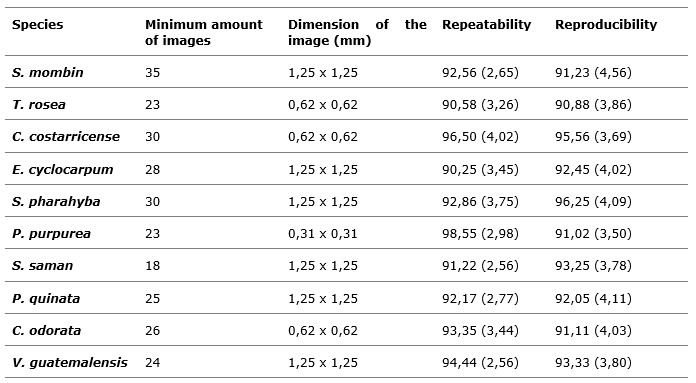
Note: Values in brackets correspond to standard deviation.
Relationship of the vessel diameter to the photographic dimension
The analysis showed a relationship between the vessel diameter and the optimal dimension of the photograph of 73 %; as the vessel is smaller, the optimal size of the photograph is lower, which influenced that small vessel species like P. purpurea could work with photographs with dimensions less than 1.25 x 1.25 mm (Figure 6). However, the vast majority of species reported that dimensions smaller than 1 mm are not functional enough to obtain statistically valid data. This was due to the loss of representativeness of the vessel data, hence species with larger vessels required a larger image. Reinig et al., (2018) found similar correlation results, determining that the size of the images had a 60 % correlation with respect to the size of the vessels, finding that in very small images in many cases the vessel is cut out, generating underestimates in its measurement; if the image is very small (less than 0.5 mm) there is a risk of having images made up entirely of vessels, which means that the representativeness of the image is null.
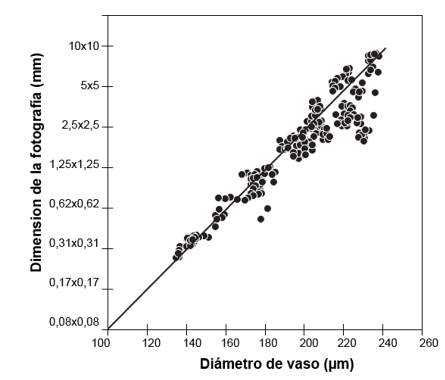
Figure 6. - Relationship between vessel diameter and photographic dimensions and
macroscopic images
of wood as a function of vessel diameter for ten timber species in Costa Rica
CONCLUSIONS
The minimum quantity of photographs per species was determined from 19 to 35, with Spondias mombin being the species with the highest requirement of photographs with 35 and Caryocar costarricense the species with the lowest quantity of photographs with 19. With respect to the optimal common dimension of photographs it was 1.25 x 1.25 mm, with Peltogyne purpurea being the species that could be worked with smaller dimensions of 0.31 x 0.31 mm; the combination of the minimum quantity of photographs and the optimal dimension allowed to be representative and reproducible in more than 90 %. Finally, a correlation was found between the size of the image and the diameter of the vessels, and it was found that, depending on the diameter of the vessels, it is possible to work with smaller photographs with a ratio of more than 70 %.
REFERENCES
ANDRADE BUENO, I. G., PICOLI, E. A. DE T., ISAIAS, R. M. DOS S., BARBOSA LOPES-MATTOS, K. L., CRUZ, C. D., NAOMI KUKI, K., & VALVERDE ZAUZA, E. A. 2020. Wood anatomy of field grown eucalypt genotypes exhibiting differential dieback and water deficit tolerance. Current Plant Biology, 22.[Consulta 1° junio 2020] Disponible en: https://doi.org/10.1016/j.cpb.2020.100136
ARZAC, A., LÓPEZ-CEPERO, J. M., BABUSHKINA, E. A., & GOMEZ, S. 2018. Applying methods of hard tissues preparation for wood anatomy: Imaging polished samples embedded in polymethylmethacrylate. Dendrochronologia, 51, 76-81. [Consulta 1° junio 2020] Disponible en: https://doi.org/10.1016/j.dendro.2018.08.005
AYALA-USMA, D. A., LOZANO-GUTIÉRREZ, R. E., & GONZÁLEZ ARANGO, C. 2019. Wood anatomy of two species of the genus Chrysochlamys (Clusiaceae: Clusioideae: Clusieae) from the northern Andes of Colombia. Heliyon, vol 5 no.7, 17. [Consulta 1° junio 2020] Disponible en: https://doi.org/10.1016/j.heliyon.2019.e02078
BEYER, R., LETORT, V., & COURNÈDE, P. H. 2014. Modeling tree crown dynamics with 3D partial differential equations. Frontiers in Plant Science, 5(JUL), 18. [Consulta 1° junio 2020] Disponible en: https://doi.org/10.3389/fpls.2014.00329
BIONDI, F., & ROSSI, S. 2015. Plant-water relationships in the Great Basin Desert of North America derived from Pinus monophylla hourly dendrometer records. International Journal of Biometeorology, vol 59 no. 8, 939-953. [Consulta 1° junio 2020] Disponible en: https://doi.org/10.1007/s00484-014-0907-4
BROWN, H. R. (2013). The Theory of the Rise of Sap in Trees: Some Historical and Conceptual Remarks. Physics in Perspective, vol 15 no. 3, 320-358. [Consulta 1° junio 2020] Disponible en: https://doi.org/10.1007/s00016-013-0117-1
BROWN, L. A., OGUTU, B. O., & DASH, J. 2020. Tracking forest biophysical properties with automated digital repeat photography: A fisheye perspective using digital hemispherical photography from below the canopy. Agricultural and Forest Meteorology, 287, 107-944. [Consulta 1° junio 2020] Disponible en: https://doi.org/10.1016/j.agrformet.2020.107944
BROWNSON, K., ANDERSON, E. P., FERREIRA, S., WENGER, S., FOWLER, L., & GERMAN, L. 2020. Governance of Payments for Ecosystem Ecosystem services influences social and environmental outcomes in Costa Rica. Ecological Economics, 174, 106-659. [Consulta 1° junio 2020] Disponible en: https://doi.org/10.1016/j.ecolecon.2020.106659
CAMPELO, F., GUTIÉRREZ, E., RIBAS, M., SÁNCHEZ-SALGUERO, R., NABAIS, C., & CAMARERO, J. J. 2018. The facultative bimodal growth pattern in Quercus ilex A simple model to predict sub-seasonal and inter-annual growth. Dendrochronologia, 49 (March), 77-88. [Consulta 1° junio 2020] Disponible en: https://doi.org/10.1016/j.dendro.2018.03.001
CARLSSON, J., & ISAKSSON, P. 2018. Dynamic crack propagation in wood fibre composites analysed by high speed photography and a dynamic phase field model. International Journal of Solids and Structures, 144-145, 78-85. [Consulta 1° junio 2020] Disponible en: https://doi.org/10.1016/j.ijsolstr.2018.04.015
COPINI, P., DECUYPER, M., SASS-KLAASSEN, U., GÄRTNER, H., MOHREN, F., & DEN OUDEN, J. (2015). Effects of experimental stem burial on radial growth and wood anatomy of pedunculate oak. Dendrochronologia, 33, 54-60. [Consulta 1° junio 2020] Disponible en: https://doi.org/10.1016/j.dendro.2014.12.001
DANTAS, L. A., & CUNHA, A. 2020. An integrative debate on learning styles and the learning process. Social Sciences & Humanities Open, vol 2 no.1, 100-017. [Consulta 1° junio 2020] Disponible en: https://doi.org/10.1016/j.ssaho.2020.100017
PAZ PÉREZ-OLVERA, C., & DÁVALOS-SOTELO, R. (2008). Algunas características anatómicas y tecnológicas de la madera de 24 especies de Quercus (encinos) de México. Madera Bosques, vol 14no.3, 43-80. [Consulta 1° junio 2020] Disponible en: https://doi.org/10.21829/myb.2008.1431206
MICCO, V., ARONNE, G., & BAAS, P. 2008. Wood anatomy and hydraulic architecture of stems and twigs of some Mediterranean trees and shrubs along a mesic-xeric gradient. Trees - Structure and Function, vol 22 no.5, 643-655. [Consulta 1° junio 2020] Disponible en: https://doi.org/10.1007/s00468-008-0222-y
SOUSA FERNANDES, D. D., DE ALMEIDA, V. E., FONTES, M. M., DE ARAÚJO, M. C. U., VÉRAS, G., & DINIZ, P. H. G. D. 2019. Simultaneous identification of the wood types in aged cachaças and their adulterations with wood extracts using digital images and SPA-LDA. Food Chemistry, 273, 77-84. [Consulta 1° junio 2020] Disponible en: https://doi.org/10.1016/j.foodchem.2018.02.035
FIORAVANTI, M., DI GIULIO, G., & SIGNORINI, G. 2017. A non-invasive approach to identifying wood species in historical musical instruments. Journal of Cultural Heritage, 27, S70-S77. [Consulta 1° junio 2020] Disponible en: https://doi.org/10.1016/j.culher.2016.05.012
GIACHI, G., CAPRETTI, C., LAZZERI, S., SOZZI, L., PACI, S., MARIOTTI LIPPI, M., & MACCHIONI, N. 2017. Identification of wood from Roman ships found in the docking site of Pisa (Italy). Journal of Cultural Heritage, 23, 176-184. [Consulta 1° junio 2020] Disponible en: https://doi.org/10.1016/j.culher.2016.07.005
GLABASNIA, A., & HOFMANN, T. 2006. Sensory-directed identification of taste-active ellagitannins in American (Quercus alba L.) and European oak wood (Quercus robur L.) and quantitative analysis in bourbon whiskey and oak-matured red wines. Journal of Agricultural and Food Chemistry, vol 54 no. 9, 3380-3390. [Consulta 1° junio 2020] Disponible en: https://doi.org/10.1021/jf052617b
HAVINGA, I., HEIN, L., VEGA-ARAYA, M., & LANGUILLAUME, A. 2020. Spatial quantification to examine the effectiveness of payments for ecosystem services: A case study of Costa Rica's Pago de Servicios Ambientales. Ecological Indicators, 108, 105-766. [Consulta 1° junio 2020] Disponible en: https://doi.org/10.1016/j.ecolind.2019.105766
KOBAYASHI, K., HWANG, S. W., OKOCHI, T., LEE, W. H., & SUGIYAMA, J. 2019. Non-destructive method for wood identification using conventional X-ray computed tomography data. Journal of Cultural Heritage, 38, 88-93. [Consulta 1° junio 2020] Disponible en: https://doi.org/10.1016/j.culher.2019.02.001
MA, F., & HUANG, A. MIN. 2018. Rapid identification and quantification three chicken-wing woods of Millettia leucantha, Millettia laurentii and Cassia siamea by FT-IR and 2DCOS-IR. Journal of Molecular Structure, 11-66, 164-168. [Consulta 1° junio 2020] Disponible en:https://doi.org/10.1016/j.molstruc.2018.04.037
MACHADO, J. S., PEREIRA, F., & QUILHÓ, T. 2019. Assessment of old timber members: Importance of wood species identification and direct tensile test information. Construction and Building Materials, 207, 651-660. [Consulta 1° junio 2020] Disponible en: https://doi.org/10.1016/j.conbuildmat.2019.02.168
MALIK, I., PAWLIK, £., ŒLÊZAK, A., & WISTUBA, M. 2019. A study of the wood anatomy of Picea abies roots and their role in biomechanical weathering of rock cracks. Catena, 173, 264-275. [Consulta 1° junio 2020] Disponible en: https://doi.org/10.1016/j.catena.2018.10.018
MARCELO-PEÑA, J. L., SANTINI, L., & TOMAZELLO FILHO, M. 2019. Wood anatomy and growth rate of seasonally dry tropical forest trees in the Marañón River Valley, northern Peru. Dendrochronologia, 55, 135-145. [Consulta 1° junio 2020] Disponible en: https://doi.org/10.1016/j.dendro.2019.04.008
PACHECO, A., CAMARERO, J. J., POMPA-GARCÍA, M., BATTIPAGLIA, G., VOLTAS, J., & CARRER, M. 2020. Growth, wood anatomy and stable isotopes show species-specific couplings in three Mexican conifers inhabiting drought-prone areas. Science of the Total Environment, 698, 134055. [Consulta 1° junio 2020] Disponible en: https://doi.org/10.1016/j.scitotenv.2019.134055
REINIG, F., GÄRTNER, H., CRIVELLARO, A., NIEVERGELT, D., PAULY, M., SCHWEINGRUBER, F., SOOKDEO, A., WACKER, L., & BÜNTGEN, U. (2018). Introducing anatomical techniques to subfossil wood. Dendrochronologia, 52, 146-151. [Consulta 1° junio 2020] Disponible en: https://doi.org/10.1016/j.dendro.2018.10.005
SÁNCHEZ-COSTA, E., POYATOS, R., & SABATÉ, S. 2015. Contrasting growth and water use strategies in four co-occurring Mediterranean tree species revealed by concurrent measurements of sap flow and stem diameter variations. Agricultural and Forest Meteorology, 207, 2437. [Consulta 1° junio 2020] Disponible en: https://doi.org/10.1016/j.agrformet.2015.03.012
SEVANTO, S., NIKINMAA, E., RIIKONEN, A., DALEY, M., PETTIJOHN, J. C., MIKKELSEN, T. N., PHILLIPS, N., & HOLBROOK, N. M. 2008. Linking xylem diameter variations with sap flow measurements. Plant and Soil, 305(12), 77-90. [Consulta 1° junio 2020] Disponible en: https://doi.org/10.1007/s11104-008-9566-8
STEPANOVA, A. V., OSKOLSKI, A. A., TILNEY, P. M., & VAN WYK, B. E. 2013. Wood anatomy of the tribe Podalyrieae (Fabaceae, Papilionoideae): Diversity and evolutionary trends. South African Journal of Botany, 89, 244-256. [Consulta 1° junio 2020] Disponible en: https://doi.org/10.1016/j.sajb.2013.07.023
Conflict of interests:
The authors declare not to have any interest conflicts.
Authors' contribution:
The authors have participated in the writing of the work and analysis of the
documents.
![]()
This work is licensed under a Creative Commons Attribution-NonCommercial 4.0
International license
Copyright (c) 2020 Juan Carlos Valverde, Dagoberto Arias, Erick Mata, Geovanni
Figueroa, Nelson Zamora