Revista Cubana de Ciencias Forestales. 2020; mayo-agosto 8(2):
296-315![]()
Estado nutrimental de Juniperus flaccida Schltdl. y Phoradendron juniperinum Engelm. en respuesta a fertilización química al suelo
Nutrient status of Juniperus flaccida Schltdl. and Phoradendron juniperinum Engelm. in response to chemical fertilization to soil
Estado nutricional de Juniperus flaccida Schltdl. E Phoradendron juniperinum Engelm. Em resposta à fertilização química do solo
Fanny Libertad
González-Torralva1* ![]() https://orcid.org/0000-0002-0708-8426
https://orcid.org/0000-0002-0708-8426
Miguel Ángel
López-López1 ![]() https://orcid.org/0000-0001-5741-8350
https://orcid.org/0000-0001-5741-8350
Marcos Jiménez-Casas2 ![]() https://orcid.org/0000-0003-0727-017X
https://orcid.org/0000-0003-0727-017X
Dionicio Alvarado-Rosales1![]() https://orcid.org/0000-0001-5941-2446
https://orcid.org/0000-0001-5941-2446
1Universidad Autónoma de Chapingo, Colegio de Postgraduados, México.
2Universidad Nacional Autónoma de México, Colegio de Postgraudados, México.
*Autor para la correspondencia: gonzalez.fanny@colpos.mx
Recibido: 6 de febrero de 2020.
Aprobado: 28 de mayo de 2020.
RESUMEN
Juniperus flaccida Schltdl. es una especie forestal muy importante que, en El Cardonal, Hidalgo, México es fuertemente atacada por Phoradendron juniperinum Engelm. El objetivo de esta investigación fue evaluar la respuesta de ramillas de J. fláccida en cuanto a su biomasa, su estado nutrimental y el de su planta parásita, con relación a las aplicaciones de nitrógeno (N), fósforo (P) y potasio (K), en un bosque natural de Juniperus flaccida. Las dosis y fuentes de fertilizantes fueron 0 y 1000; 0 y 300; y 0, 450 y 900 g árbol-1 de urea; ácido fosfórico y sulfato de potasio, respectivamente. El diseño experimental utilizado fue completamente al azar con arreglo factorial 2X2X3, con seis repeticiones. Los análisis de suelo mostraron bajos niveles de N y P, y suficientes de K en el sitio. Las aplicaciones individuales de N y P no tuvieron efectos significativos en biomasa, a pesar de ser deficientes en el área experimental. J. flaccida presentó mayores concentraciones foliares de N, Mg, Cu, Mn y B que P. juniperinum, y éste presentó mayores concentraciones de P, K, Ca, Fe y Zn que su hospedero. La aplicación de nitrógeno disminuyó significativamente la concentración de K en P. juniperinum e influyó en la partición de P foliar entre J. flaccida y P. juniperinum, en favor de la hospedera y en detrimento de la parásita. El aporte de K incrementó la concentración de N del follaje de J. flaccida. Los análisis de vectores mostraron que la combinación de fósforo con la dosis alta de potasio propició un incremento en biomasa de J. flaccida.
Palabras clave: Análisis de suelos; Análisis de vectores; Fertilizantes; Planta parásita.
ABSTRACT
Juniperus flaccida Schltdl. is a very important forest species which is severely affected by Phoradendron juniperinum Engelm. at El Cardonal, Hidalgo, Mexico. The objective of this research was to evaluate the response, in terms of tree twig biomass and nutrient status of both the tree and its parasitic species, to fertilization with nitrogen (N), phosphorus (P), and potassium (K), in a natural forest of J. flaccida. The fertilizer rates and sources were 0, 1000; 0, 300 and 0, 450, 900 g tree-1 of urea, phosphoric acid and potassium sulphate, respectively. The experimental design was a completely randomized factorial (2X2X3) with six replicates. Soil analysis showed low levels of N and P, and adequate levels of K in the site. Nitrogen or phosphorus alone did not significantly affect twig biomass, despite of being deficient nutrients in the study site. J. flaccida displayed higher concentrations of N, Mg, Cu, Mn, and B than P. juniperinum and the latter, showed higher concentrations of P, K, Ca, Fe, and Zn than its host species. Application of nitrogen significantly decreased the concentration of K in P. juniperinum and influenced the partition of foliar P between J. flaccida and P. juniperinum, in favor of the host tree and to the detriment of the parasitic species. Application of K increased foliar N in J. flaccida. Vector analysis showed that the combination of phosphorus with the highest dose of potassium led to an increase in twig biomass of J. flaccida.
Keywords: Soil analysis; Vector analysis; Fertilizers; Parasitic plant.
RESUMO
Juniperus flaccida Schltdl. É uma espécie florestal muito importante que, em El Cardonal, Hidalgo, México, é fortemente atacada por Phoradendron juniperinum Engelm. O objectivo desta investigação foi avaliar a resposta dos ramais de J. flaccida em termos da sua biomassa, do seu estado nutricional e do da sua planta parasita, em relação às aplicações de nitrogénio (N), fósforo (P) e potássio (K), numa floresta natural de Juniperus flaccida. As doses e fontes de fertilizantes foram 0 e 1000; 0 e 300; e 0, 450 e 900 g de ureia de árvore 1; ácido fosfórico e sulfato de potássio, respectivamente. O desenho experimental utilizado foi completamente aleatório num arranjo fatorial 2X2X3, com seis réplicas. As análises do solo mostraram níveis baixos de N e P, e K suficiente no local. As aplicações individuais de N e P não tiveram um efeito significativo na biomassa, apesar de serem deficientes na área experimental. J. flaccida apresentou concentrações foliares de N, Mg, Cu, Mn e B mais elevadas do que P. juniperinum, e este último apresentou concentrações mais elevadas de P, K, Ca, Fe e Zn do que o seu hospedeiro. A aplicação de azoto diminuiu significativamente a concentração de K em P. juniperinum e influenciou a divisão do foliar P entre J. flaccida e P. juniperinum, em favor do hospedeiro e em detrimento do parasita. A contribuição de K aumentou a concentração de N da folhagem de J. flaccida. As análises vectoriais mostraram que a combinação do fósforo com a elevada dose de potássio levou a um aumento da biomassa de J. flaccida.
Palavras-chave: Análise dos solos; Análise de vetores; Fertilizantes; Planta parasita.
INTRODUCCIÓN
Las perturbaciones bióticas afectan una amplia gama de especies arbóreas en todos los climas y contribuyen a aumentar las tasas de mortalidad de los árboles a nivel mundial (Szmidla et al., 2019). El muérdago pertenece a un grupo extenso de plantas parásitas que establece relaciones duraderas con una amplia gama de especies de árboles hospederos. Con el cambio climático, el estrés ecofisiológico está aumentando, lo que puede promover que los árboles sean más susceptibles a la infección por muérdago, pudiendo incluso conducir hacia mayores tasas de mortalidad en las masas forestales (Griebel et al., 2017).
Phoradendron juniperinun (muérgado de táscate) se encuentra actualmente parasitando a Juniperus flaccida (táscate) en la región de El Cardonal, en el Estado de Hidalgo, México; según la literatura no existen registros de distribución de esta especie de muérdago en el área de estudio (Geils et al., 2002), lo que significa que se está extendiendo al encontrar condiciones adecuadas para su establecimiento. Esta interacción entre P. juniperinum y J. flaccida está causando graves perturbaciones al bosque natural de J. flaccida en la región. Dentro de las interacciones fisiológicas más importantes entre plantas hospederas y plantas huésped, se incluye las relaciones nutrimentales e hídricas (Glatzel y Geils 2009). No obstante, la importancia de tales relaciones entre árboles hospederos y muérdagos, se conoce relativamente poco, especialmente en lo referente a requerimientos nutrimentales y su abastecimiento en las plantas parásitas (Okubamichael et al., 2011; Raya-Pérez et al., 2014).
Velasco (1999), menciona que llevar a cabo un manejo nutrimental de los árboles a través de la fertilización química, representa una actividad importante en el control de plagas y enfermedades, así como un componente integral en la productividad ya sea en el área agrícola o forestal. Cuando los árboles están establecidos en suelos pobres o con deficiencia de algún nutrimento, responderán mostrando un deficiente crecimiento y desarrollo, así como susceptibilidad al ataque de patógenos. Según Reyes-Pozo et al., (2018), las plantas cuyas condiciones nutrimentales son balanceadas se vuelven más tolerantes o menos susceptibles ante el ataque de plagas y/o enfermedades puesto que pueden protegerse a posibles infecciones y/o a limitar las presentes.
El estado nutrimental y vigor de los muérdagos depende en gran medida del estado nutrimental de los hospederos, de los que absorbe los nutrimentos, por lo que una cabal comprensión de las relaciones nutrimentales entre hospedero y huésped puede conducir a derivar posibles estrategias de manejo nutrimental de los árboles para disminuir su susceptibilidad al ataque por plantas parásitas. Bajo este contexto, el presente estudio pretendió evaluar la respuesta nutrimental de J. flaccida y P. juniperinum, su planta parásita en la región, a la fertilización química del hospedero con N, P y K de manera individual y combinada.
MATERIALES Y MÉTODOS
El experimento se realizó de agosto 2018 a septiembre 2019, en la localidad de San Miguel Tlazintla, El Cardonal, Hidalgo, México, en un bosque natural de Juniperus flaccida Schltdl. El predio se encuentra en las coordenadas 18° 42' 20,2" latitud norte y 90° 44' 09,0" longitud oeste y a una altitud de 2 094 m. El sitio presenta un rango de temperatura entre 12 22°C y una precipitación pluvial media de 430 mm. Cuenta con un clima semi-seco templado con lluvias en verano; el suelo dominante es leptosol (INEGI 2009) (Figura 1).
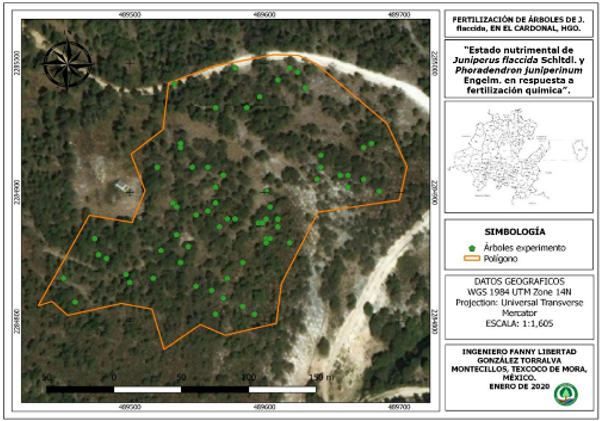
Figura 1. - Área de estudio; Bienes Comunales San Miguel Tlazintla, El Cardonal, Hidalgo, México
Diseño experimental y de tratamientos
Se realizó un experimento factorial 2x2x3, con seis repeticiones, tomando como unidad experimental un árbol. La Tabla 1, muestra los factores (nutrimentos) de variación que fueron probados, tales como, nitrógeno (N), fósforo (P) y potasio (K). Los factores estudiados se definieron con base en un análisis químico de suelo realizado previamente, mientras que los niveles (dosis de fertilización) de cada uno de ellos se determinó con base en los resultados publicados de un ensayo de fertilización en una plantación de 12 años de edad de Cupressus lusitanica en Colombia (Tschinkel, 1972).
Tabla 1.- Factores de variación del experimento factorial de fertilización establecido
en el predio ejidal
de San Miguel Tlazintla, Municipio de El Cardonal, Hidalgo
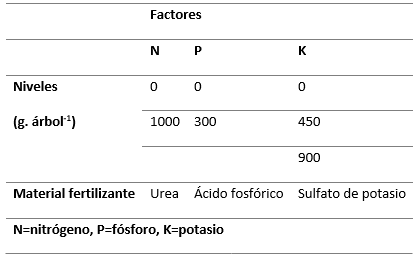
La combinación de los niveles de los factores estudiados produjo 12 tratamientos que se mencionan a continuación, cuyos valores entre paréntesis corresponden a las cantidades aplicadas (g.árbol-1) de urea, ácido fosfórico y sulfato de potasio, en ese orden: T1 (0,0,0); T2 (0,0,450); T3 (0,0,900); T4 (0,300.0); T5 (0,300,450); T6 (0,300,900); T7 (1000,0,0); T8 (1000,0,450); T9 (1000,0,900); T10 (1000,300,0); T11 (1000,300,450) y T12 (1000,300,900).
Fertilización
La aplicación de tratamientos se realizó empezando con la selección de las unidades experimentales, procurando que éstas fueran homogéneas en cuanto a diámetro, a fin de evitar efectos por diferencias de tamaño. Además, se procuró que todas las unidades experimentales tuvieran el mismo grado de incidencia de muérdagos. Seleccionados los árboles, se realizó la medición del Diámetro a la Altura del Pecho (DAP) de cada individuo con la ayuda de una cinta diamétrica, se le aplicó el tratamiento de fertilización de acuerdo a una aleatorización previamente realizada y se etiquetó. El DAP se midió solamente para asegurar uniformidad de las dimensiones de los árboles experimentales.
Los fertilizantes se aplicaron en forma granulada para el caso de nitrógeno (urea) y potasio (sulfato de potasio), en el caso de fósforo (ácido fosfórico) se aplicó de forma líquida mezclando la dosis determinada en cinco litros de agua y vertiendo la solución sobre el área de goteo, con una regadera.
Variables evaluadas
Las variables evaluadas fueron la fertilidad del suelo (análisis químicos), potencial hídrico para ambas especies; variable de crecimiento (incremento en biomasa de ramillas de J. flaccida) y variables fisiológicas (concentración nutrimental en el follaje de J. flaccida y P. juniperinum).
El potencial hídrico fue tomado con la ayuda de una cámara de presión de Scholander, modelo 3005 Soil Moisture. Para ello, se seleccionaron cuatro individuos de J. flaccida y en cada uno de ellos se seleccionó una rama. Dentro de cada rama seleccionada, se recolectó una ramilla madura y libre de daños físicos o por agentes bióticos, cercana a una planta de P. juniperinum, de la que también se recolectó una ramilla. El corte de las muestras se realizó de manera uniforme recolectando una ramilla de 10 cm aproximadamente; posteriormente ésta se introdujo en la cámara de presión, abriendo la válvula de gas y tomando la medición en el momento en que una burbuja de savia emergió de la base de la ramilla.
La obtención de biomasa de ramillas se tomó retirando con el uso de una tijera telescópica, cinco ramillas del último flujo de crecimiento. Éstas se distinguieron por la coloración clara que presentaron. Las muestras fueron trasladadas al laboratorio donde se secaron en una estufa de secado marca FELISA a 70ºc durante 72 horas. Una vez secas las muestras se pesaron en una balanza analítica, obteniendo así la biomasa de las ramillas de cada uno de los árboles pertenecientes al experimento de fertilización.
El estado nutrimental inicial se realizó a partir de una recolecta de follaje de ambas especies en nueve árboles seleccionados aleatoriamente dentro del área experimental, siguiendo el protocolo de Wells y Allen (1985). Las muestras fueron procesadas y enviadas al laboratorio de Nutrición Vegetal del Colegio de Postgraduados, para la determinación de las concentraciones foliares de N, P, K, Ca, Mg, Fe, Cu, Zn, Mn y B.
Doce meses después de establecido el experimento de fertilización, se llevó a cabo una toma de muestras de follaje de tres individuos por tratamiento, las cuales se obtuvieron del tercio superior de la copa, mismas que fueron analizadas para la determinación de N, P y K, con la finalidad de conocer la respuesta de los árboles a la aplicación de los tratamientos.
Durante el muestreo de suelos realizado antes de la aplicación de los tratamientos, se tomaron ocho muestras distribuidas al azar dentro del área de estudio, a una profundidad de 30 cm. A partir de estas muestras, se formaron cuatro muestras compuestas, las cuales fueron llevadas al mismo laboratorio que las muestras foliares, para la determinación de pH, MO, CIC (Capacidad de Intercambio Catiónico), N, P, K Ca, Mg, Zn, Fe, Mn y Cu. Los resultados de suelos obtenidos se interpretaron con la ayuda de la NOM-021 SEMARNAT 2001 y fueron la base para decidir los nutrimentos a probar en el experimento.
Métodos de análisis químico
La determinación de las concentraciones de P y K se realizó en el extracto resultante de la digestión húmeda de material vegetal seco con ácido sulfúrico, ácido perclórico y peróxido de hidrógeno (1,3:0,7:1, v:v:v), de acuerdo con lo descrito por Alcántar y Sandoval (1999). Los extractos fueron leídos en un equipo de espectrometría de emisión e inducción por plasma (Agilent 725 Series ICP-OES).
La concentración de N se determinó en una alícuota de 10 ml del digestado antes descrito, mediante destilación de la muestra y valoración mediante titulación con ácido sulfúrico.
Análisis estadístico
Con la ayuda del paquete estadístico SAS (versión 9.0), se llevó a cabo el análisis estadístico que consistió en la realización de un Análisis de Varianza (ANAVA) y una prueba de comparación de medias de Tukey (α=0,05), para los efectos de tratamientos sobre la concentración de N, P y K en el follaje de J. flaccida y P. juniperinum después de los tratamientos de fertilización. La biomasa de ramillas, junto con las concentraciones y contenidos nutrimentales se analizó mediante la técnica de análisis gráfico de vectores (Timmer y Stone, 1978). Mediante análisis de regresión, se examinó la dependencia de las concentraciones foliares de P. juniperinum con respecto a las de Juniperus fláccida. Los potenciales hídricos de J. fláccida y P. juniperinum se compararon mediante una prueba de T.
RESULTADOS Y DISCUSIÓN
Análisis de suelo y foliares preliminares
De acuerdo con la Tabla 2, los suelos del sitio de estudio, interpretados de acuerdo con la Norma Oficial Mexicana 021 de la SEMARNAT (Secretaría de Medio Ambiente y Recursos Naturales 2002) sólo presentan niveles bajos de nitrógeno y fósforo, mientras las demás propiedades analizadas se encuentran en niveles adecuados o ligeramente altos.
Tabla 2. - Propiedades químicas del suelo en el bosque de Juniperus flaccida del predio ejidal de San Miguel Tlazintla, El Cardonal, Hidalgo
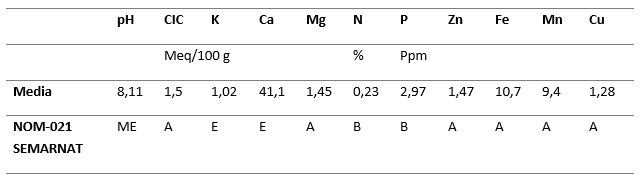
ME: Medianamente Alcalino; A: Adecuado; B: Bajo; E: Elevado.
Los resultados de los análisis foliares preliminares (antes de la aplicación de los tratamientos), en lo relativo a los macroelementos (N, P, K, Ca y Mg) se muestran en la Figura 2; mientras que los referentes a las concentraciones de los microelementos (Fe, Cu, Zn, Mn y B) se presentan en la Figura 3.
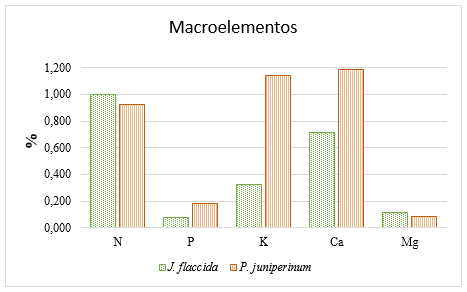
Figura 2. - Concentraciones de N, P, K, Ca y Mg en el follaje de J. flaccida y P. juniperinum,
antes de
la aplicación de los tratamientos de fertilización
La Figura 2, muestra que los muérdagos presentan altas concentraciones de P (0,182 %), K (1,146 %) y Ca (1,188 %), comparadas con las correspondientes concentraciones en J. flaccida. Estos resultados concuerdan con los obtenidos por Raya-Pérez et al., 2014, quienes observaron que los muérdagos (Psittacanthus calyculatus) acumularon niveles altos de potasio, nutrimento que en la presente investigación está 253 % por encima de la concentración que presenta el hospedero (J. flaccida). Por otro lado, Glatzel (1983), investigó las concentraciones de otros nutrimentos y observó que el N fue el único nutrimento no acumulado por los muérdagos en niveles más altos que sus hospederos, coincidiendo con lo obtenido en los análisis reflejados en la Figura 2, donde J. flaccida presenta una concentración foliar de N superior en 8,78 %, que la concentración de este nutrimento en P. juniperinum. Como en el caso del N, en el presente estudio, la concentración de Mg en el hospedero fue superior en 34 % respecto a la concentración del mismo nutrimento en la planta huésped.
Los análisis obtenidos en la presente investigación concuerdan con un estudio realizado por Panvini y Eickmeier (1993), quienes obtuvieron las concentraciones minerales de Phoradendron leucarpum y de varias especies de hospederos, encontrando que el muérdago tuvo concentraciones nutrimentales de 0,97 a 2,88 veces mayores que sus árboles hospederos. En dicho estudio los datos recopilados mostraron concentraciones superiores de K, N, Ca, Mg, P, Na, Fe, Zn y Cu en el huésped, comparados con los del hospedero.
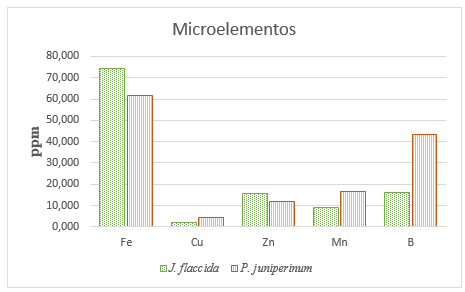
Figura 3. - Concentraciones de Fe, Cu, Zn, Mn y B en el follaje de J. flaccida y P. juniperinum,
antes de
la aplicación de tratamientos de fertilización
En la Figura 3 se presentan las concentraciones de microelementos y se observan mayores concentraciones de Cu (4,47 ppm), Mn (16,81 ppm) y B (43,35 ppm), en P. juniperinum comparadas con las encontradas en J. fláccida (1,98, 9,11 y 16,11 ppm, respectivamente). Estos microelementos se encuentran 66, 46 y 63 % en menor concentración en la especie hospedera que en la planta parásita.
Es muy probable que las mayores concentraciones de la mayoría de nutrimentos encontradas en el muérdago, con respecto a las del hospedero, sean una adaptación relacionada con la necesidad del huésped de absorber agua desde el hospedero (Raftoyannis et al., 2015); es decir, la planta huésped (muérdago) necesita absorber agua del xilema del árbol hospedero. Dado que el agua siempre se mueve de mayores potenciales hídricos hacia sitios con menores potenciales hídricos, para que el muérdago pueda absorber agua desde el árbol hospedero, se requiere que los potenciales hídricos del muérdago sean menores (más negativos) que aquellos del hospedero. Las altas concentraciones de sales en los tejidos del muérdago contribuyen a lograr esa condición, con lo que se genera un flujo de agua desde el árbol (en este caso J. flaccida), hacia el muérdago (P. juniperinum).
Las determinaciones de potenciales hídricos (Ψ) en ramillas del árbol hospedero y de la planta huésped indicaron que el Ψ medio en P. juniperinum (-36.12 bar) es significativamente menor (Pr>0,009; Tabla 3) que el de J. fláccida (Ψ=-29.75 bar). Estos resultados corroboran lo que se ha obtenido en otras investigaciones en donde se ha demostrado que los muérdagos mantienen tasas de transpiración más altas y son menos eficientes en el uso de agua que sus hospederos (Garkoti et al., 2002). Hosseini et al., (2007), observaron que Viscum album, especie parásita de Carpinus betulus, mantiene una tasa de transpiración más alta, con la finalidad de lograr la absorción principalmente de nitrógeno. Por otro lado, Glatzel y Geils (2009), mencionan que los muérdagos mantienen un potencial hídrico más negativo que las especies hospederas, con la finalidad de evitar el cierre estomatal y marchitamiento, lo que puede explicar los resultados obtenidos en la presente investigación, en la que se corrobora el bajo potencial hídrico que mantiene P. juniperinum con respecto a J. flaccida.
Tabla 3. - Prueba de t para el potencial hídrico de Juniperus flaccida y Phoradendron
juniperinum
en el Ejido San Miguel Tlazintla, El Cardonal, Hidalgo, México

G.L.: Grados de libertad
Estado nutrimental de J. flaccida y P.
juniperinum
En las Figuras 4 a, b y c se presentan las concentraciones foliares de N, P y K, respectivamente para las especies de interés. En el caso del nitrógeno (Figura 4a), en todos los tratamientos J. flaccida (especie hospedera) presentó las mayores concentraciones en comparación con P. juniperinum (especie huésped), destacando los tratamientos T6 (P 300-K 900) y T12 (N 1000-P 300-K 900), los cuales presentaron las mayores concentraciones de este nutrimento en J. flaccida.
De acuerdo con la Figura 4a, en todos los tratamientos, incluyendo aquellos que no contemplaron la aplicación de nitrógeno, la concentración foliar de este nutrimento en el huésped se encuentra, en promedio, 18 % por debajo de la concentración del mismo nutrimento en el árbol hospedero. Estos resultados concuerdan con los obtenidos por Bowie y Ward (2004), quienes determinaron que el N es el único nutrimento que se encontró en menores concentraciones en el muérdago (65 %), condición que concuerda con la teoría de la absorción pasiva de nutrimentos.
Los resultados obtenidos en la presente investigación difieren con lo obtenido por Türe et al., (2010), puesto que los análisis realizados por dichos autores mostraron que el nitrógeno se almacenó en una mayor concentración en el muérdago Viscum álbum, que en las especies hospederas bajo observación. Esto significa que las proporciones entre las concentraciones de N en plantas hospederas y huéspedes pueden ser características asociadas a la propia combinación de especies hospederas y huéspedes.
Panvini y Eickmeier (1993) sugieren que el N es un nutrimento que limita el crecimiento del muérdago y que el mecanismo que utilizan para poder acceder a este nutrimento es mantener altas tasas de transpiración. Lo anterior coincide con los resultados del presente estudio, que demuestran que los muérdagos mantienen potenciales hídricos más bajos que la especie hospedera.
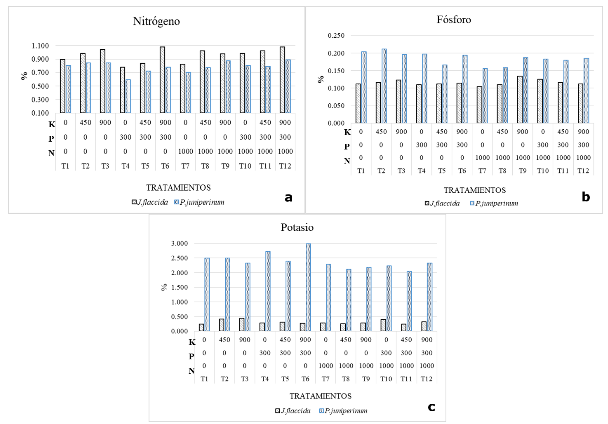
Figura 4.- Concentración foliar de N, P y K en J. flaccida y P. juniperinum, en un experimento de
fertilización
en predio ejidal de San Miguel Tlazintla, municipio de El Cardonal, Hidalgo
En la Figura 4b se observan las concentraciones de fósforo para los doce tratamientos en ambas especies. J. flaccida presenta en promedio, 33 % menor concentración de fósforo que P. juniperinum. Dentro de los tratamientos analizados, destacan el segundo (450 g de K) y el primer tratamiento (testigo), los cuales presentan una mayor concentración de P, aun cuando este nutrimento no fue suministrado a los árboles de esos tratamientos. La aplicación de P a los árboles no se manifestó en incrementos de las concentraciones de este nutrimento en el follaje de ninguna de las dos especies (J. fláccida o P. juniperinum; Figura 4b), probablemente debido a que el P aplicado se convirtió en fosfato de calcio, como consecuencia de las elevadas condiciones de alcalinidad del sitio (Hopkins y Ellsworth 2005); sin embargo, la Figura 4b muestra claramente que la aplicación de N sí afectó la repartición del P entre el hospedero y el huésped, advirtiéndose que la aplicación de 1000 g de urea por árbol favoreció la acumulación de P en el hospedero, en detrimento del P foliar de la planta parásita (Tabla 4).
No existen referencias en la literatura, relacionadas con el efecto anteriormente señalado; sin embargo, este hallazgo sugiere que la aplicación de N en áreas infestadas similares al sitio estudiado, pudiera conducir a mejorar la partición de P entre el hospedero y el huésped, en favor del primero, lo cual pudiera, a la postre, mejorar el vigor del árbol, especialmente en sitios con deficiencia de P. Los resultados de la presente investigación, concuerdan con los obtenidos por autores como Glatzel (1983), Panvini y Eickmeier (1993), Bowie y Ward (2004) y Türe et al., (2010), quienes encontraron que el P, es uno de los nutrimentos que se almacena en mayor concentración en el tejido del muérdago en comparación con las especies hospederas analizadas.
Tabla 4. - Efectos de la aplicación de nitrógeno sobre la partición de N, P y K foliares,
entre Juniperus flaccida y Phoradendron juniperinum
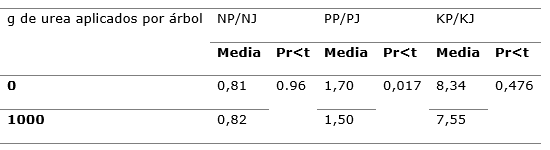
NP: Concentración foliar de N en P.
juniperinum; NJ: Concentración foliar de N en Junuperus flaccida; PP: Concentración foliar de P en P. juniperinum; PJ:
Concentración foliar de P en J. flaccida; KP: Concentración foliar de K en P. juniperinum; KJ: Concentración foliar de K en J. flaccida.
El potasio (Figura 4c) es uno de los nutrimentos principales en escenarios de árboles invadidos por muérdagos, el cual se almacena en muy elevadas concentraciones, dentro de la planta parásita (Glatzel 1983; Türe et al., 2010 y Tennakoon et al., 2011). Los autores mencionados sostienen que la mayoría de especies vegetales tienen preferencia por la absorción de este nutrimento, puesto que ejerce un papel como transportador de otros elementos, además de que es esencial para la osmorregulación y el control estomático.
El análisis de varianza de la Tabla 5 muestra los efectos de los tratamientos de fertilización en las concentraciones de nitrógeno, fósforo y potasio en el follaje de J. flaccida y P. juniperinum.
Los análisis de varianza indican que las concentraciones foliares de N, P y K fueron afectadas por los tratamientos (Tabla 5). Se observa que el N tuvo efectos significativos sobre las concentraciones de fósforo y potasio en el follaje de P. juniperinum (Pr = 0,03 y Pr = 0,01, respectivamente). Por otro lado, el K, influyó sobre la concentración de nitrógeno en el follaje de J. flaccida (Pr=0,01).
En el caso de las interacciones entre nutrimentos, no se encontraron efectos significativos sobre las concentraciones de N, P y K en el follaje de ninguna de las especies. Sin embargo, destaca la interacción de N*P, que presentó una ligera influencia sobre las concentraciones de N en J. flaccida (Pr=0,07) y de P en P. juniperinum (Pr=0,07; Tabla 5).
Tabla 5. - Significancia según los análisis de varianza, de los efectos de tratamientos de fertilización sobre
las concentraciones de N, P y K en el follaje
de J. flaccida y P. juniperinum, en el predio ejidal de San
Miguel Tlazintla, Municipio de El Cardonal, Hidalgo
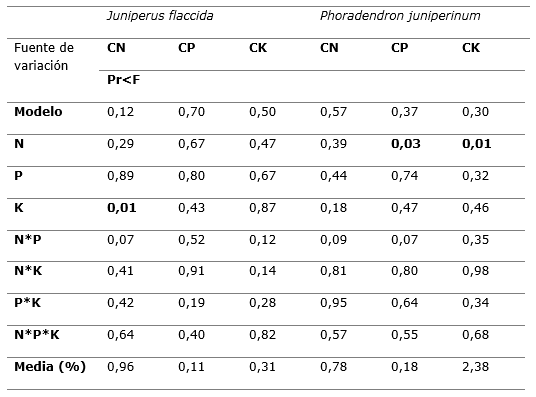
Pr<F (α=0,05); N: Nitrógeno; P: Fósforo; K: Potasio; CN: Concentración foliar de nitrógeno;
CP: Concentración foliar de fósforo; CK: Concentración foliar de potasio.
La comparación de medias de Tukey (α = 0,05, Tabla 6), muestra que la aplicación de fósforo no tuvo efectos significativos sobre las concentraciones de N, P y K en el follaje de ninguna de las dos especies de estudio; sin embargo, al aplicar N a los árboles, se presentó una media menor en la concentración de potasio en el follaje de P. juniperinum, lo que sugiere que ésta práctica puede contribuir a reducir el vigor del muérdago, al disminuir la concentración de solutos en el individuo, dificultando el flujo de agua y iones desde el árbol hospedero. Por otro lado, se observa que, con la aplicación de 900 g de K, se presentó un aumento significativo en la concentración foliar de N en el follaje J. flaccida (Tabla 6). Este efecto pudiera tener implicaciones sobre las decisiones de manejo nutrimental de los bosques de J. flaccida afectados por muérdagos, toda vez que el abastecimiento de K al árbol hospedero mejora la nutrición de N, a la vez que la aplicación de N disminuye la absorción de K por parte de la planta parásita.
Tabla 6. - Prueba de Tukey
(α=0,05) para las concentraciones de N, P y K en el follaje de J. flaccida y
P. juniperinum,
después de los tratamientos de fertilización
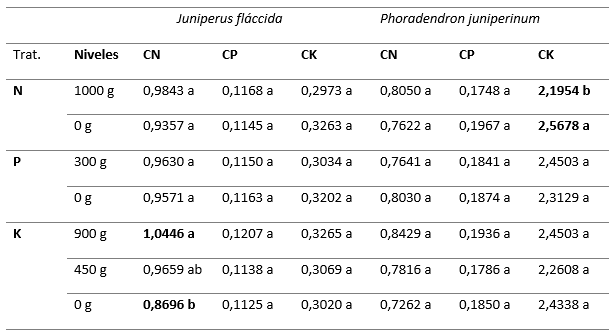
N: Nitrógeno; P: Fósforo; K: Potasio; CN: Concentración foliar de Nitrógeno; CP: concentración foliar
de fósforo; CK:
concentración foliar de potasio; dentro de un tratamiento y un nutrimento foliar, medias con
la misma letra no son estadísticamente diferentes
(α=0,05).
En la Figura 5 (a, b y c) se muestra el comportamiento que tuvo el incremento en biomasa, concentración y contenido de N, P y K en el follaje de J. flaccida, con respecto a los tratamientos de fertilización.
El tratamiento P*K2 (300 g P * 900 g K por árbol) presentó los mayores crecimientos en términos de biomasa de cinco ramillas de J. flaccida, comparado con el resto de los tratamientos (Figura 5). Esto coincide con los datos de diámetros obtenidos para los árboles que recibieron este tratamiento, mismos que presentaron mayores incrementos en diámetro o evitaron el decremento de esta variable que en otros tratamientos causó la sequía que se presentó en el área de estudio durante el año experimental (2018).
En el caso del N (Figura 5a) se observa que tres de los doce tratamientos (P*K1, N y P) presentan concentraciones de nitrógeno más bajas que la concentración crítica (0.826 %) generada para J. fláccida (González-Torralva 2020). La falta de respuesta de los demás tratamientos se debió quizá a algún otro factor limitante, puesto que algunos tratamientos como N*P*K2 (1000 g N, 300 g P y 900 g K), N*K1 (1000 g N y 450 g K), K2 (900 g K), N*P*K1 (1000 g N, 300 g P y 450 g K) y N*P (1000 g N y 300 g P), generaron altas concentraciones de N, pero no se obtuvo respuesta del incremento en biomasa de ramillas. Esto quizá pudo deberse a la condición de sequía a la que estuvo sujeta el área experimental, o a algún otro factor limitante. En cuanto a los tratamientos en donde se combinó N*K, la poca respuesta pudo deberse quizá al antagonismo que existe entre estos nutrimentos (Mengel y Kirkby 1982).
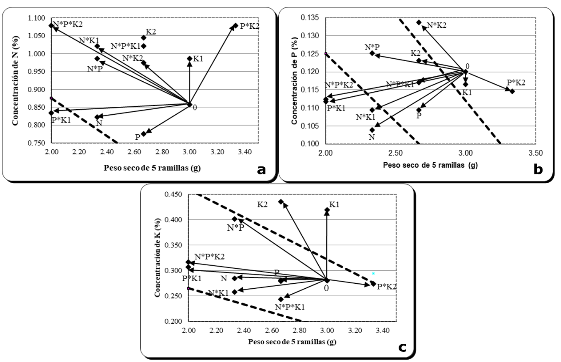
Figura 5. - Efecto de tratamientos sobre la biomasa de cinco ramillas, concentración
y contenido
de nutrimentos en un experimento de fertilización con N, P y K en un bosque natural de J. flaccida, en
El Cardonal, Hidalgo
En cuanto al P, el análisis de suelo indicó deficiencia de este elemento. De acuerdo con esto, al agregar fósforo se esperaría mayor crecimiento de los árboles; sin embargo, esto no sucedió de manera significativa. En la Figura 5b, se observa que el tratamiento P*K2 (300 g P * 900 g K por árbol), presentó un considerable incremento en biomasa de ramillas (3.33 g), comparado con el resto de los tratamientos, los cuales presentaron biomasa incluso por debajo del tratamiento testigo, así como disminución en la absorción de fósforo, aun cuando este nutrimento se aplicó. Una posible explicación a este comportamiento es que el suelo dentro del área de estudio presenta altos niveles de calcio, que, al realizar la aplicación de fósforo, contribuye a formar fosfatos de calcio, lo que vuelve indisponible al fósforo para la planta (Mengel y Kirkby 1982). Este tratamiento (P*K2) presenta una concentración de P (0,115 %) muy cercana a la concentración crítica (0,11 %) reportada para esta especie (González-Torralva, 2020).
En lo relativo a potasio, el análisis de suelos en el área de estudio indica que existe un nivel elevado de este nutrimento, por lo que se esperaría que, al realizar la aplicación de K, no se obtuviera respuesta de los individuos en cuanto a crecimiento. En la Figura 5c, se observa que el mejor tratamiento en términos de incremento en biomasa de cinco ramillas es P*K2 (300 g P * 900 g K), pero resultó bajo en concentración de K, puesto que presenta 0,274 % de K en el follaje de J. flaccida, una concentración menor a la crítica reportada para esta especie (0,301 %; González-Torralva, 2020). En el caso de los tratamientos K1 y K2, (450 y 900 g de K, respectivamente), su incremento en biomasa se ubicó incluso por debajo del correspondiente al tratamiento testigo; especialmente el tratamiento K2, pero la concentración de K para ambos tratamientos fue casi de 0.100 % por encima de la concentración crítica de la especie de interés. Estas altas concentraciones de K en el follaje y bajo crecimiento son acordes con el diagnóstico de suficiencia de K en el sitio (análisis de suelo), pero son contrarias al mayor crecimiento en diámetro, registrado en los árboles que recibieron K, en cuyo caso, es probable que el abastecimiento de K haya contribuido a la conservación de la turgencia de las células del fuste del árbol, evitando así la reducción de diámetro que experimentaron los árboles que no recibieron este tratamiento. Vázquez-Cisneros et al., (2018) reportaron que una plantación experimental de Pinus greggii, creciendo en suelo con suficiencia de potasio, mostró respuestas positivas en crecimiento cuando se aplicó este nutrimento, concluyendo que el potasio posiblemente mejoró la turgencia de las células en las plantas y la incorporación de carbono.
Al realizar una integración de los resultados de análisis de suelo, foliares y de crecimiento de los árboles se puede afirmar que, en el sitio, a pesar de que el análisis de suelo indica altos niveles de potasio, al realizar la aplicación de este nutrimento se obtuvieron los mejores diámetros.
Relaciones funcionales entre las concentraciones foliares de N, P y K entre J. flaccida y P. juniperinum
La Figura 6 (a, b y c) muestra el comportamiento de la concentración foliar de nutrimentos en Phoradendron juniperinum, en función de las correspondientes concentraciones nutrimentales en Juniperus flaccida.
La relación entre el N foliar en P. juniperinum y el correspondiente en J. flaccida (Figura 6a) es descrita por una curva de saturación (R2=0,77) que indica que cuando la concentración foliar de N en J. fláccida es inferior a 0,975 %, la concentración del mismo nutrimento en el follaje de P. juniperinum aumenta conforme lo hace la de J. flaccida; sin embargo, cuando la concentración de N en el árbol hospedero alcanza la mencionada concentración de N, la planta parásita se vuelve incapaz de acumular más nitrógeno en su follaje. Este comportamiento puede ser relevante desde el punto de vista del manejo nutrimental y control de la planta parásita, pues sugiere que la aplicación de N es una práctica que puede ayudar a manejar el estado de este nutrimento en el árbol, aun en presencia de la planta parásita, con el efecto adicional de hacer disminuir la concentración de K en el muérdago (Tabla 6). El valor de R2 indica que 77 % de la concentración de nitrógeno presente en el follaje del muérdago depende de la concentración nutrimental del hospedero. Eso explica que cuando hay poco nitrógeno en los tejidos de J. flaccida, conforme aumenta la concentración de N en esta especie, aumenta también en el muérdago, pero cuando el hospedero tiene abundante N, este nutrimento probablemente se vuelve suficiente para la planta parásita y las concentraciones de nitrógeno en ésta se tornan más variables debido posiblemente a que aparecen otros factores limitantes del crecimiento de la misma.
En el caso de P (Figura 6b), el comportamiento es similar al de nitrógeno, sólo que, en este caso, el modelo que mejor representó la relación entre el P foliar del árbol y el del muérdago fue de tipo lineal, con una R2 de 0,1106. Es decir, al incrementar la concentración foliar de P en el hospedero, incrementa la concentración del nutrimento en el muérdago, aunque en este caso, la concentración foliar de P en J. flaccida solamente explica un 11 % de la concentración de P en el muérdago. Esto significa que la absorción de P por parte del muérdago se ve influenciada por un mayor número de factores y es más compleja que en el caso del N.
La Figura 6c, muestra que cuando las concentraciones de K en el follaje de J. flaccida son bajas, en el muérdago pueden ser bajas o altas, pero cuando tales concentraciones en el hospedero son altas, aparentemente en el muérdago son bajas; si esto es consistente, entonces indica que en esta región del estado de Hidalgo, la aplicación de K al suelo elevaría las concentraciones de éste nutrimento en J. flaccida sin generar mayores aumentos en la concentración del mismo en el muérdago, lo que podría contribuir a aumentar los diferenciales de potenciales hídricos entre el hospedero y el parásito, a favor del primero (menores potenciales en J. flaccida que en P. juniperinum), pudiendo dificultar la absorción de agua y otros nutrimentos por parte del muérdago y promover su debilitamiento.
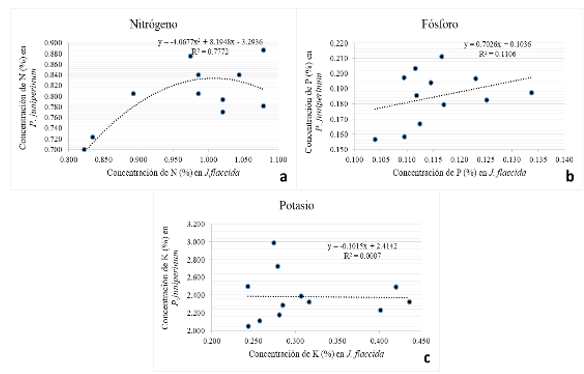
Figura 6. - Comportamiento de la concentración de nutrimentos (N, P y K) en P.
juniperinum en función
de las correspondientes concentraciones en J. flaccida
Anteriormente se hizo mención que el muérdago tiene muy elevados requerimientos de K, por lo que, al disminuir la concentración de este nutrimento en el tejido de la planta parásita, se generaría un desbalance nutrimental, lo que pudiera disminuir el vigor de los muérdagos y aumentar el vigor de los árboles. En el presente estudio, este comportamiento se reflejó en que los árboles que recibieron K presentaron un incremento en diámetro o al menos se evitó un decremento del mismo por condiciones de sequía, como sucedió en árboles que no recibieron este nutrimento.
El P, K, Ca, Fe y Zn registran mayores concentraciones en follaje de muérdago que en el de Juniperus fláccida; lo contrario ocurre en los casos de N y Mg, Cu, Mn y B.
El potencial hídrico es inferior en ramillas de P. juniperinum que en ramillas de J. flaccida localizadas cerca de la base del muérdago.
La aplicación de N influye en la partición de P foliar entre J. flaccida y P. juniperinum, en favor de la especie hospedera y en detrimento de la especie parásita.
La aplicación de N disminuye significativamente la concentración foliar de K en la planta parásita, mientras que el suministro de K incrementa la concentración de N en J. flaccida.
La correlación entre la concentración foliar de N en J. flaccida y la correspondiente en P. juniperinum se pierde cuando las concentraciones de N en J. flaccida son elevadas.
No existe correlación entre la concentración foliar de K en J. flaccida y la correspondiente concentración en P. juniperinum.
La combinación de P con la dosis alta de K es el único tratamiento que promueve el incremento en biomasa de ramillas en J. fláccida en el área de estudio.
AGRADECIMIENTOS
Al Consejo Nacional de Ciencia y Tecnología (CONACYT), por la beca otorgada al primer autor para el desarrollo de sus estudios de Maestría en Ciencias.
Al Técnico Forestal, Ing. Eduardo Vargas Solís y al Comisariado Ejidal C. Alfredo Isidro Cervantes, por haber permitido llevar a cabo la investigación en sus terrenos ejidales.
REFERENCIAS BIBLIOGRÁFICAS
ALCÁNTAR, G. G. y SANDOVAL, M. V. 1999. Manual de análisis químico de tejido vegetal. Guía de muestreo, preparación, análisis e interpretación. Publicación especial No. 10 de la Sociedad Mexicana de la Ciencia del Suelo A. C. Chapingo, México. 156 p.
BOWIE, M., y WARD, D. 2004. Water and nutrient status of the mistletoe Plicosepalus acaciae parasitic on isolated Negev Desert populations of Acacia raddiana differing in level of mortality. Journal of Arid Environments, 56(3), 487508. Recuperado de: https://doi.org/10.1016/S0140-1963(03)00067-3
GARKOTI, S.C., AKOIJAM, S.B. y SINGH. S.P. 2002. Ecology of water relations between mistletoe (Taxillus vestitus) y its host oak (Quercus floribunda). Tropical Ecology 43: 243-249. Recuperado de: http://docplayer.net/35453400-Ecology-of-water-relations-between-mistletoe-taxillus-vestitus-and -its-host-oak-quercus-floribunda.html
GEILS, B. W.; CIBRIÁN, T. J.; MOODY, B. 2002. Mistletoes of North American Conifers. Gen. Tech. Rep. RMRSGTR98. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station. 123 p.
GLATZEL, G. 1983. Mineral nutrition and water relations of hemiparasitic mistletoes: a question of partitioning. Experiments withLoranthus europaeus onQuercus petraea and Quercus robur. Oecologia, 58(1983), pp.310-32. Recuperado de: https://doi.org/10.1007/BF00379691
GLATZEL, G. y GEILS, B.W. 2009. Mistletoe ecophysiology: host-parasite interactions. Botany 87: 10-15. Recuperado de: https://doi.org/10.1139/B08-096
GONZÁLEZ-TORRALVA. F. L. 2020. Estándares Nutrimentales, Fertilización Química de Juniperus flaccida Schltdl. y nutrición de su huésped parásito Phoradendron juniperinum Engelm. Tesis de Maestría en Ciencias. Colegio de Postgraduados, Texcoco, México. 80 pp.
GRIEBEL, A.; WATSON, D. y PENDALL, E. 2017. Mistletoe, friend and foe: synthesizing ecosystem implications of mistletoe infection. Environmental Research Letters 12: 9 pp. Recuperado de: https://doi.org/10.1088/1748-9326/aa8fff
HOPKINS, B. y ELLSWORTH J. 2005. Phosphorus Availability with Alkaline/Calcareous Soil. Western Nutrient Management Conference Proceedings. Vol. 6. Salt Lake City, UT, USA. pp. 88-93. Recuperado de: https://www.researchgate.net/publication /237734366_Phosphorus_availability_with_alkalinecalcareous_soil
HOSSEINI, S.M.; KARTOOLINEJAD, D.; MIRNIA, S.K.; TABIBZADEH, Z.; AKBARINIA, M. y SHAYANMEHR, F. 2007. The effects of Viscum album L. on foliar weight and nutrient content of host trees in Caspian forests [Iran]. Recuperado de: http://desert.semnan.ac.ir/uploads/kartoli_article7.pdf
INEGI. Instituto Nacional de Estadística Geografía e Informática, 2009. Prontuario de información geográfica municipal de los Estados Unidos Mexicanos. Cardonal, Hidalgo. Clave Geoestadística 13015.
MENGEL, K. y KIRKBY A. E. 1982. Principles of plant nutrition. International Potash Institute. Berna, Suiza. 593 p.
SECRTETARÍA DE MEDIO AMBIENTE Y RECURSOS NATURALES (SEMARNAT). 2002. NOM-021-RECNAT-2000 Que establece las especificaciones de fertilidad, sanidad y clasificación de suelos, estudios, muestreo y análisis. Diario Oficial de la Federación. 31 de diciembre del 2002. 85 p.
OKUBAMICHAEL, D. Y.; GRIFFITHS, M. E. y WARD, D. 2011. Host specificity, nutrient and water dynamics of the mistletoe Viscum rotundifolium and its potential host species in the Kalahari of South Africa. Journal of Arid Environments, 75(10), 898902. Recuperado de: https://doi.org/10.1016/j.jaridenv.2011.04.026
PANVINI, A. D. y EICKMEIER, W. G. 1993. Nutrient and water relations of the mistletoe Phoradendron leucarpum (Viscaceae): how tightly are they integrated? American Journal of Botany, 80 (1993), pp. 872-878. Recuperado de: http://doi.wiley.com/10.1002/j.1537-2197.1993.tb15307.x
RAFTOYANNIS, Y.; RADOGLOU, K.; BREDEMEIER, M. 2015. Effects of mistletoe infestation on the decline and mortality of Abies cephalonica in Greece. Annals ofForest Research 58(1): 1-11. DOI: 10.15287/afr.2015.347. Recuperado de: https://www.afrjournal.org/index.php/afr/article/view/347
RAYA-PÉREZ, J. C.; RAMÍREZ-PIMENTEL, J. G.; COVARRUBIAS-PRIETO, J.; ACEVEDO-LARA, B. y AGUIRRE-MANCILLA, C. 2014. Mineral and Chlorophyll Content of the Psittacanthus calyculatus (DC) G. Don Hemiparasitic Plant and Four Host Trees. Revista Chapingo Serie Ciencias Forestales y del Ambiente, XX (1), 109-117. Recuperado de: http://dx.doi.org/10.5154/r.rchscfa.2013.06.017
REYES-POZO, J. L.; LEÓN-SÁNCHEZ, M. A. y HERRERO-ECHEVERRIA. G. 2018. Efecto de la Fertilización Sobre el Volumen de Madera en Pinus caribaea Morelet var. caribaea Barret y Golfari en Cub. Foresta Veracruzana, 20(2), 29. Recuperado de: https://www.redalyc.org/jatsRepo/497/49758340003/html /index.html
SZMIDLA, H.; TKACZYK, M.; PLEWA, R.; TARWACKI, G. Y SIEROTA, Z. 2019. Impact of common mistletoe (Viscum album L.) on scots pine forests A call for action. Forests 10 (847):1-15. doi:10.3390/f10100847 Recuperado de: https://www.mdpi.com/1999-4907/10/10/847
TENNAKOON K. U.; CHAK W. H. y BOLIN J. F. 2011. Nutritional and isotopic relationships of selected Bornean tropical mistletoehost associations in Brunei Darussalam. Functional Plant Biology 38: 505513. Recuperado de: https://doi.org/10.1071/FP10211
TIMMER, V. y STONE, E. 1978. Comparative Foliar Analysis of Young Balsam Fir Fertilized With Nitrogen, Phosphorus, Potassium, and Lime 1. Soil Science Society of America Journal, Vol. 42, 125-130 pp. Recuperado de: doi:10.2136/sssaj1978.03615995004200010027x
TSCHINKEL, H. 1972. Factores Limitantes del Crecimiento de Plantaciones de Cupressus lusitánica en Antioquia, Colombia. Revista Facultad Nacional de Agronomía Medellín, 27(2), 3-55. Recuperado de: https://revistas.unal.edu.co/index.php/refame/article/view/29961
TÜRE, C.; BÖCÜK, H. y AªAN, Z. 2010. Nutritional relationships between hemi-parasitic mistletoe and some of its deciduous hosts in different habitats. Biologia, 65(5). Recuperado de: https://doi.org/10.2478/s11756-010-0088-5
VÁZQUEZ-CISNEROS, I., PRIETO-RUIZ, J. Á., LÓPEZ-LÓPEZ, M. Á., WEHENKEL, C.,
DOMÍNGUEZ-CALLEROS, P. A. y MUÑOZ-SÁEZ, F. E. 2018. Crecimiento y supervivencia de una plantación de Pinus greggii Engelm. ex Parl. var.
greggii bajo diferentes tratamientos de fertilización. Revista Chapingo Serie Ciencias Forestales y del
Ambiente 24(2), 251-264. doi: 10.5154/r.rchscfa.2017.05.036. Recuperado de:
https://www.researchgate.net/publication/323970315_Crecimiento_y_supervivencia_de_una_plantacion_de_Pinus_greggii_Engelm_ex_Parl_var_greggii_bajo_diferentes_
tratamientos_de_fertilizacion
VELASCO V. A. 1999. Papel de la nutrición mineral en la tolerancia a las enfermedades de las plantas. Terra Latinoamericana. 17 (3). 192-200. Recuperado de: https://www.redalyc.org/pdf/573/57317303.pdf
WELLS, C.G. y ALLEN, L. 1985. A Loblolly Pine Management Guide: When and Where to Apply Fertilizer (No. SE-GTR-36). U.S. Department of Agriculture, Forest Service, Southeastern Forest Experiment Station, Asheville, NC. Recuperado de: https://doi.org/10.2737/SE-GTR-36
Conflicto de intereses:
Los autores declaran no tener conflictos de intereses.
Contribución de los autores:
Los autores han participado en la redacción del trabajo y análisis de los documentos.
![]()
Esta obra está bajo una licencia de Creative Commons Reconocimiento-NoComercial
4.0 Internacional.
Copyright (c) 2020
Fanny Libertad
González-Torralva,
Miguel Ángel
López-López,
Marcos Jiménez-Casas,
Dionicio Alvarado-Rosales