Revista Cubana de Ciencias Forestales. May-August, 2019, 7(2): 225-239![]()
Translated from the original in spanish
Needles anatomical differences of Pinus tropicalis Morelet in extreme soil of slate and white sandy
Diferencias anatómicas en acículas de Pinus tropicalis Morelet en edátopos extremos de pizarras y arenas blancas en Pinar del Río
Liutmila Pérez-Del Valle1
Gretel Geada López1
Rogelio Sotolongo-Sospedra1
1Universidad de Pinar del Río "Hermanos Saíz Montes de Oca". Pinar del Río, Cuba. E-mail: delvalle@upr.edu.cu , gabriel@upr.edu.cu , soto@upr.edu.cu
Received: February 3rd, 2019.
Approved: May 5th, 2019.
ABSTRACT
P. tropicalis is a species endemic to western Cuba, prioritized in Forest Development Plans until 2030. It develops in very contrasting edatopes of white sands and slates, so over the years it must show anatomical differences that allow it to grow. The objective of the study was to characterize the anatomical variation in transversal cuts of the needles of the species in three localities of Pinar del Río. Differences were detected in the anatomical variables of the needles. The variables number of stomas, thickness of the hypodermis and number of resin channels separate San Ubaldo on white sands from the rest. The discriminant analysis shows with 87% certainty the classification of the cases in each of the collection localities (a priori) and the variables number of stomata, number of channels, cuticle thickness and thickness of hypodermis the most discriminant. Preliminarily it is inferred that soil conditions and water availability influence foliar anatomy.
Krywords: Pinus tropicalis; needle anatomy; anatomical variation.
RESUMEN
P. tropicalis es una especie endémica del occidente de Cuba, priorizada en los Planes de Desarrollo Forestal hasta el 2030. Se desarrolla en edátopos muy contrastantes de arenas blancas y pizarras, por lo que a lo largo de los años debe mostrar diferencias anatómicas que le permitan crecer. El estudio tuvo como objetivo caracterizar la variación anatómica en cortes transversales de las acículas de la especie en tres localidades de Pinar del Río. Se detectaron diferencias en las variables anatómicas de las acículas. Las variables número de estomas, grosor de la hipodermis y número de canales resiníferos logran separar la localidad San Ubaldo sobre arenas blancas de las restantes. El análisis de discriminante muestra con un 87 % de certeza la clasificación de los casos en cada una de las localidades de colecta (a priori) y las variables número de estomas, número de canales, grosor de cutícula y grosor de hipodermis las más discriminantes. Preliminarmente se infiere que las condiciones edáficas y la disponibilidad de agua influyen en la anatomía foliar.
Palabras clave: Pinus tropicalis; anatomía de las acículas; variación anatómica.
INTRODUCTION
Anatomical and morphological characterization of needles has been widely used to distinguish within-species adaptations to particular environmental conditions (Köbölkuti et al., 2017; Jankowski et al., 2017; Kivimäenpää et al, 2017) and in some cases as characters to identify or differentiate species of the same genus, phenotypically similar, particularly in Pinus (García-Álvarez et al., 2009; Iglesias et al., 2012; Dörken and Stützel, 2012; Meng et al., 2018).
Schoettle and Rochelle, (2000) in Pinus flexilis reported a low morphological variation in tree growth variables (DAP, H, length, annual and foliar increments) between sites located between 1 630 and 3 330 masl and a marked variation in anatomical variables of stomata density and the number of rows of these, associated with a high physiological plasticity or a wide ecophysiological tolerance. Reinhardt et al., (2011), in an ecophysiological study in two origins of this species, verified a significant effect of altitude on physiological variables related to gas exchange and photosynthesis and their relation with the variation in the anatomy of the stomatological apparatus.
In species with Euroasian distribution, P. sylvestris and P. uncinata (Boratyñska and Pashkevich, 2001; Boratyñska and Bobowicz, 2001), anatomical variations were found at the epidermis of the acícula that facilitated differentiation between the two species in sympatric habitats. (2003) used the morphology and anatomy of the needles to compare two populations of Pinus uliginosa and quantify the degree of hybridization of the P. mugo x P. sylvestris complex in the taxonomic definition of P. uliginosa, through variables such as number of stomas and resin channels.
García-Álvarez et al., (2009) highlight the importance of the characteristics of foliar structure in the identification and classification of species of the genus Pinus of the Iberian Mediterranean, from palynological sheets with the use of cuticle and stoma characteristics.
Other studies show that variables associated with resin channels are feasible for taxonomic definition in conifers (Dörken and Stützel, 2012; Sheue et al., 2014; Ghimire et al., 2014) and among populations of a species as an adaptive trait to altitude and exposure (Tiwari et al., 2013; Köbölkuti et al., 2017; Jankowski et al., 2017).
Ju-Kyeong and Jung-Oh, (2013) in an anatomical study of the needles in 289 individuals of Abies koriana, detected a clinal variation in number, type of resin channels, width of the needles and thickness of the needles, which would be related to adaptation to more extreme environments. In more recent studies, Èortan et al., (2017) demonstrate that stomata are genetically controlled traits, so they can be used for differentiation studies between Populus nigra populations in three valleys of the Danube river. Thus, anatomical variables such as length, width, stoma shape, pore width, stoma density, shape coefficient and stoma area showed significant differences between populations.
Pinus tropicalis Morelet is endemic to western Cuba and constitutes a relict of the Eurasian lineage in Central America and the Caribbean (Geada-López et al., 2002; Geada-López et al., 2004; Gernandt et al., 2005). The species forms continuous pure or sympathetic pine forests with Pinus caribaea Morelet var. caribaea Barret and Golfari, on very extreme edatopes: white silica sands and slates (López-Almiral, 1982 and Geada-López et al., 2004) reported morphological differences in the cone and needles and at chloroplastic DNA level between populations corresponding to edatopes of white sands and slates (López-Almiral, 1982 and Geada-López et al., 2004).
According to the aspects mentioned above and taking into account that P. tropicalis inhabits the most extreme edatopes due to its hydric and nutritional characteristics, this research aims to characterize the anatomical variation in the species' needles in three localities, differentiate the populations according to the anatomical variables and determine the variable or variables that most contribute to this differentiation. All this will provide the basis for plans for the selection of differences in seed sources that achieve better establishment and productivity depending on the site to be planted.
MATERIALS AND METHODS
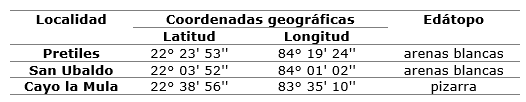
Sampling: the study was carried out in three localities corresponding to natural populations of P. trpoicalis: Petriles, San Ubaldo and Cayo la Mula in the province of Pinar del Río (Table 1). Needles were collected from the middle portion of each tree at a rate of at least 30 trees per locality.
Table 1. - Georeferencing of needle collection localities of P. tropicalis
Measured anatomical variables
The following anatomical variables were measured: number of stomas, number of channels, cuticle thickness, epidermis thickness, hypodermis thickness, chlorophyll parenchyma thickness, transfusion parenchyma thickness, distance to the right channel, distance to the left channel. Values were averaged for statistical analysis. Additionally, the type of resin channel was classified according to Sheue et al., (2014). The measurements were made with an optical microscope model NLCD-307B.
Statistical analysis
Analysis of main components: it was used with the objective of revealing the relationships between variables, in order to reduce the dimensionality of the analysis and to explain the relationships with a minimum number of dimensions and to express the combined variability of all the variables. Components with auto-values greater than one were considered; the component matrix was rotated according to the Varimax method. For this analysis, the effect of the locality factor was not taken into account.
Comparison between localities: The Kruskal-Wallis test (k independent samples) was used to establish whether there were differences between localities for each of the anatomical variables studied; and the MannWhitney U test (two independent samples), both for p = 0.05 to check whether there was a significant effect of the locality factor.
Classification of localities: the multivariate technique of discriminant analysis was used in order to maximize the differences between localities, considered these as dependent variables or classification variables. All anatomical variables were considered as independent variables. Another objective of this analysis was to identify the variables that most contribute to differentiate localities.
RESULTS AND DISCUSSION
Anatomical characteristics of the needles of P. tropicalis
Figure 1 represents the equifacial structure of a cross section of a P. tropicalis acticle. On the outside there is a cuticle defined as a modification of the uniestratified epidermal tissue. Towards the interior is located a hypodermic tissue formed by several layers of non-uniform cells; followed to these the very well defined chlorophyll parenchyma is located, forming packages delimited by the wide channels. In the interior there is the very wide parenchyma of transfusion and in the centre the vascular beams xylem and phloem (Figure 1).
Resin channels varied in number, but were classified as marginal type (Sheue et al., 2014); they may occupy various tissues, from epidermal to chlorophyll parenchyma. This marginal type character is considered ancestral to the genus (Dörken and Stützel, 2012; Seong and Offner, 2013; Sheue et al., 2014) and agrees with the basal position within the lineage of the Pinus section: Pinus subsection of this species in the molecular phylogeny (Gernandt et al., 2005; Gernandt et al., 2008); Keeley, 2012).
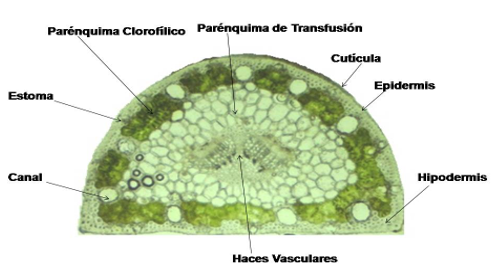
Fig. 1. - Anatomical structure of an acícula cross section of P. tropicalis in San Ubaldo
Relationship between anatomical needle structures of P. tropicalis
According to the results of the analysis of main components, the relations between the variables of the foliar anatomy of P. tropicalis can be explained by three dimensions, whose autovalues are greater than one and which express 55 % of the present variability (Table 2).
Table 2.- Analysis of main components carried out on the correlation matrix of the anatomical variables of P. tropicalis needles
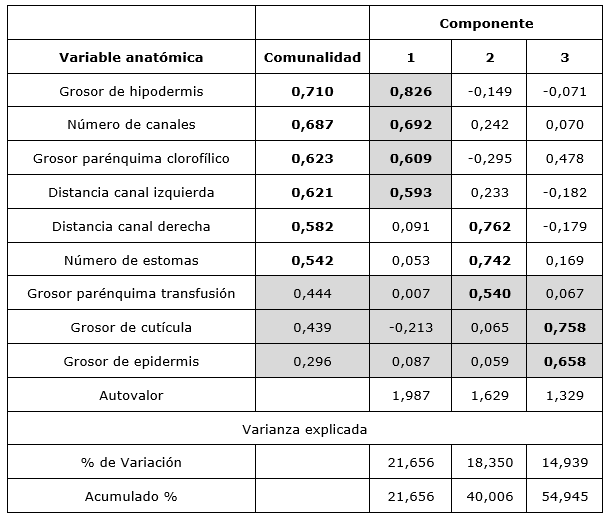
According to the value of the communities, the variables that most contributed to the segregation of the components were in order of the extraction value (greater than 0.5): thickness of hypodermis, number of channels, thickness parenchyma chlorophyll, left channel distance, right channel distance and number of stomata.
The first component explains the relationship between hypodermis thickness, number of stomas and number of channels; all three variables correlate significantly (p<0.01) and positively. The relationship between these three variables could be associated with the water environment in which the species develops.
The thickness of the hypodermis, which has up to four layers of cells, may be the result of adaptation to the control of water loss. The number of stomas, although contradictorily high for the environment where the species develops, is very cleft, sometimes reaching the chlorophyll parenchyma. This behavior has been reported by García-Álvarez et al., (2009); Tiwari et al., (2013) and Köbölkuti et al., (2017) for pines growing in very dry environments.
The second component, which explains the relationship of channel distances to the epidermis, shows the high symmetry on both sides for all localities; this coincides with that described for many phylogenetically related species where a marginal arrangement of these channels is described (Seong and Offner, 2013; Sheue et al., 2014).
Chlorophyll parenchyma thickness, although it contributes to the total variance in the analysis, is a variable that does not correlate significantly with the rest, therefore, it is not useful when explaining the relationships between them.
Behavior of anatomical variables by localities
In table 3, the average values of the variables studied by localities are presented, according to the Kruskal-Wallis test, with a significant effect of the locality factor only on the variable number of stomata (Table 3).
Table 3. - Average behaviour of the variables studied by locality
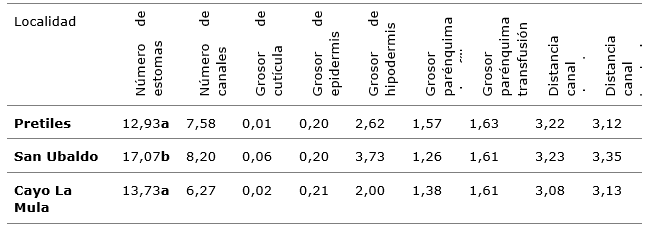
Unequal letters indicate significant differences p<0.05 MannWhitney test U
Stomatic development and differentiation are determined by genetic factors (He et al., 1998), but the morphology and anatomical types of pine needles also depend on abiotic factors (Schoettle and Rochelle, 2000); Zhang et al., 2012 and Jankowski et al., 2017).
Therefore, it can be deduced that the highest average number of stomata in the samples from the locality of San Ubaldo may be related to the soil characteristics, which are characterized by extremely sandy soils, with very little water retention, and a high light reflection due to the light color of its surface.
Jinxing et al., (2001) point out that stomach parameters are affected by factors such as altitude, atmospheric CO2 concentration (Loumala et al., 2005; Hartikainen et al., 2009), temperature, light intensity and light reflection (Kivimäenpää et al., 2017). Tiwari et al., (2013) and Köbölkuti et al., (2017) report that for both P. roxburguii and P. sylvestris stomata organization has an important role in the adjustment of plants to different altitudes, detecting that stomata density increases with elevation, due to less light scattering and a higher rate of transpiration. In contrast, Schoettle and Rochelle (2000), in the species Pinus flexilis find that in the mountains there is a significant decrease in stomata density with elevation, related to an acclimatization response to restrict water loss at high altitudes.
Although statistically not significant, both the anatomical characteristics of the P. tropicalis needles in the locality of San Ubaldo and the number of resin channels, cuticle thickness and hypodermis thickness are superior to the rest of the localities and the thickness of the chlorophyll parenchyma is lower. These characteristics may also be anatomical adjustments to the unique soil and microclimatic conditions of these ectopes.
Ju-Kyeong and Jung-Oh. (2013) and Tiwari et al., (2013), in a comparative study of Abies koreana and P.roxburguii needles, proved that the characteristics of the resin channels have a clinal and altitudinal variation. On the other hand, O'Neill, et al., (2002), in a study of the effect of geographic variation on the characteristics of resin channels in spruce seedlings, found that channel size varied between localities and was associated with summer extension, aridity index and continentality effect.
Finally, P. tropicalis has marginal channels and their number varies according to the availability of water in the localities, but more extensive studies are necessary to explain this relationship.
Classification of localities according to the anatomical characteristics of the needles
Table 4 presents the contribution of each of the variables used for classification.
Table 4. - Test of equality of means among groups
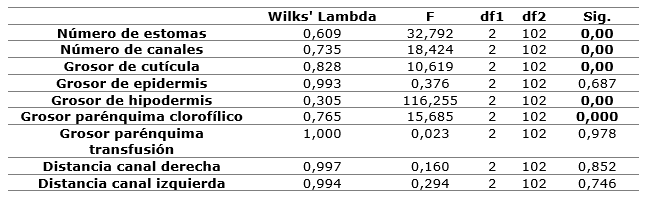
It was found that the variables number of stomas, number of channels, cuticle thickness, hypodermis thickness and chlorophyll parenchyma thickness have unequal averages in the discriminant variables, thus contributing to differentiate localities. According to the Wilks' Lambda test, the two functions obtained are capable of discriminating (p<0.05) among the groups.
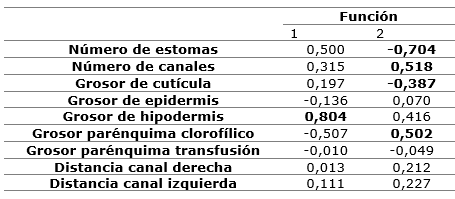
The first function achieves to explain the maximum of differences between the localities, according to the values of the centroids of the discriminant functions, in particular between the locality of San Ubaldo with respect to the other two groups (Table 5). The variable that most contributes to distinguish this group, according to the standardized values of the coefficients of the discriminant functions, is the thickness of the hypodermis, followed by the number of stomas and channels. The second function discriminates the two remaining localities, which are characterized by fewer stomas, fewer channels, less cuticle thickness and greater thickness of chlorophyll parenchyma with respect to San Ubaldo (Table 6).
Due to the importance of the number of channels for discrimination, it is valid to point out that intraspecific variations have been reported in Picea mexicana (Flores-López et al., 2013), associated with conditions of higher humidity and lower aridity, and in Picea sitchensis and Picea glauca associated with a decrease due to the effect of continentality in populations within their distribution area (O'Neill et al., 2002). Similarly, Tiwari et al., (2013) found variations along a latitudinal gradient with a decrease in these structures. Jankowski et al., (2017), in a study of P. sylvestris obtained that cold adaptation brought with it variations in anatomy, demonstrating that there is a positive relationship between temperature, number and volume of channels.
However, for an insular pine with a small distribution areal such as P. tropicalis this is the first time that a marked difference is found in the anatomy of the acícula.
Table 5. - Centroides of discriminant functions by groups (locality)
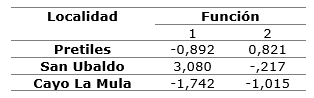
Table 6. - Standardized coefficients of canonical discriminant functions
In figure 2, the centroids of each one of the groups or localities and the location are represented, according to the values obtained from the discriminant functions, of each one of the cases. A greater grouping is observed in the localities of Pretiles and Cayo la Mula, which also have around 20 % overlap; that is, cases of one or another locality that present characteristics of the other (Table 7).
This corroborates the singularity from the edaphic point of view of the locality of San Ubaldo, which, as mentioned above, must determine the anatomical characteristics that prevail in this locality (Figure 2).
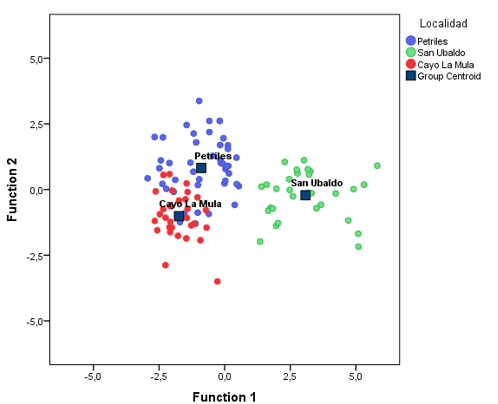
Fig. 2 - Distribution of cases and centroids in each locality based on the first two discriminant functions
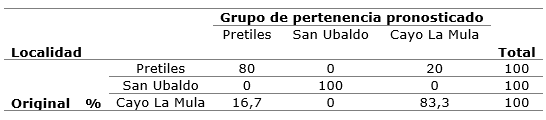
Discriminant functions classify 100 % of the cases in the locality of San Ubaldo, clearly distinguishing it from the rest of the localities. Table 6 presents a summary of the analysis, through which 86.7% of the cases were correctly classified according to the original group. Therefore, it is possible to classify, based on anatomical variables, localities; this could indicate that, in certain populations, there was a certain degree of adaptation or acclimatization to the environment.
Table 7. - Classification results
Several researchers have recognized that the analysis of morphological characteristics must be carried out in a well-conceived situation, which means that collections must be throughout the entire range of the species.
The anatomy of the P. tropicalis needles corresponds to a semicircular equifacial structure with large marginal resin channels, cleft stomata, transfusion parenchyma and well-defined chlorophyll.
There is a great variation in the anatomical variables between localities and a significant relationship between the variables thickness of hypodermis, number of stomas and number of channels, aspect that could be related to the hydric conditions where the species develops.
The anatomical variables number of stomas, number of canals, cuticle thickness and hypodermis thickness correctly classify the three localities.
BIBLIOGRAPHICAL REFERENCES
BORATYÑSKA, K. y BOBOWICZ, M.A., 2001. Pinus uncinata Ramond taxonomy based on needle characters. Plant Systematics and Evolution [en línea], vol. 227, no. 3, pp. 183-194. [Consulta: 6 mayo 2019]. ISSN 1615-6110. DOI 10.1007/s006060170047. Disponible en: https://doi.org/10.1007/s006060170047.
BORATYÑSKA, K., BORATYÑSKI, A. y LEWANDOWSKI, A., 2003. Morphology of Pinus uliginosa (Pinaceae) needles from populations exposed to and isolated from the direct influence of Pinus sylvestris. Botanical Journal of the Linnean Society [en línea], vol. 142, no. 1, pp. 83-91. [Consulta: 6 mayo 2019]. ISSN 0024-4074. DOI 10.1046/j.1095-8339.2003.00156.x. Disponible en: https://academic.oup.com/botlinnean/article/142/1/83/2433528.
BORATYÑSKA, K. y PASHKEVICH, N.A., 2001. Variability in needle traits of Pinus mugo Turra in the Ukrainian Carpathians. Acta Societatis Botanicorum Poloniae [en línea], vol. 70, no. 3, pp. 181-186. [Consulta: 6 mayo 2019]. ISSN 2083-9480. DOI 10.5586/asbp.2001.023. Disponible en: https://pbsociety.org.pl/journals/index.php/asbp/article/view/asbp.2001.023.
CORTAN, D., VILOTIC, D., SIJACIC-NIKOLIC, M. y MILJKOVIC, D., 2017. Leaf stomatal traits variation within and among black poplar native populations in Serbia. Bosque (Valdivia) [en línea], vol. 38, no. 2, pp. 337-345. [Consulta: 6 mayo 2019]. ISSN 0717-9200. DOI 10.4067/S0717-92002017000200011. Disponible en: https://scielo.conicyt.cl/scielo.php?script=sci_abstract&pid=S0717-92002017000200011&lng=es&nrm=iso&tlng=en.
DÖRKEN, V.M. y STÜTZEL, T., 2012. Morphology, anatomy and vasculature of leaves in Pinus (Pinaceae) and its evolutionary meaning. Flora - Morphology, Distribution, Functional Ecology of Plants [en línea], vol. 207, no. 1, pp. 57-62. [Consulta: 6 mayo 2019]. ISSN 0367-2530. DOI 10.1016/j.flora.2011.10.004. Disponible en: http://www.sciencedirect.com/science/article/pii/S0367253011001563.
EO, J.-K. y HYUN, J.-O., 2013. Comparative anatomy of the needles of Abies koreana and its related species. Turkish Journal of Botany [en línea], vol. 37, pp. 553-560. DOI 10.3906/bot-1201-32. Disponible en: https://www.researchgate.net/publication/296923510_Comparative_anatomy_of_the_needles_of_Abies_koreana_and_its_related_species.
FLORES-LÓPEZ, C.; GEADA-LÓPEZ, G.; LÓPEZ-UPTON, J. 2013. Morphological variation in natural population of Picea mexicana Martínez. Revista Forestal Baracoa, 32(2):3-9.
GARCÍA ÁLVAREZ, S., GARCÍA AMORENA, I., RUBIALES, J.M. y MORLA, C., 2009. The value of leaf cuticle characteristics in the identification and classification of Iberian Mediterranean members of the genus Pinus. Botanical Journal of the Linnean Society [en línea], vol. 161, no. 4, pp. 436-448. [Consulta: 6 mayo 2019]. ISSN 1095-8339. DOI 10.1111/j.1095-8339.2009.01011.x. Disponible en: https://onlinelibrary.wiley.com/doi/abs/10.1111/j.1095-8339.2009.01011.x.
GEADA LÓPEZ, G., KAMIYA, K., HARADA, K., 2004. PHYLOGENY OF DIPLOXYLON PINES (SUBGENUS PINUS). Forest Genetics [en línea], vol. 11, pp. 213-221. Disponible en: https://www.researchgate.net/publication/270902683_PHYLOGENY_OF_DIPLOXYLON_PINES_SUBGENUS_PINUS.
GERNANDT, D.S., LÓPEZ, G.G., GARCÍA, S.O. y LISTON, A., 2005. Phylogeny and Classification of Pinus. Taxon [en línea], vol. 54, no. 1, pp. 29-42. [Consulta: 6 mayo 2019]. ISSN 0040-0262. DOI 10.2307/25065300. Disponible en: https://www.jstor.org/stable/25065300.JSTOR
GERNANDT, D.S., MAGALLÓN, S., GEADA LÓPEZ, G., ZERÓN FLORES, O., WILLYARD, A. y LISTON, A., 2008. Use of Simultaneous Analyses to Guide Fossil Based Calibrations of Pinaceae Phylogeny. International Journal of Plant Sciences [en línea], vol. 169, no. 8, pp. 1086-1099. [Consulta: 6 mayo 2019]. ISSN 1058-5893. DOI 10.1086/590472. Disponible en: https://www.jstor.org/stable/10.1086/590472.
GHIMIRE, B., LEE, C. y HEO, K., 2014. Leaf anatomy and its implications for phylogenetic relationships in Taxaceae s. l. Journal of Plant Research, vol. 127, no. 3, pp. 373-388. ISSN 1618-0860. DOI 10.1007/s10265-014-0625-3.
HARTIKAINEN, K., NERG, A.-M., KIVIMÄENPÄÄ, M., KONTUNEN-SOPPELA, S., MÄENPÄÄ, M., OKSANEN, E., ROUSI, M. y HOLOPAINEN, T., 2009. Emissions of volatile organic compounds and leaf structural characteristics of European aspen (Populus tremula) grown under elevated ozone and temperature. Tree Physiology, vol. 29, no. 9, pp. 1163-1173. ISSN 0829-318X. DOI 10.1093/treephys/tpp033.
He, X. Q.; Lin, Y.H.; Lin, J.X. 1998. Research on correlation between stomatal density and variation of atmospheric carbon dioxide during a century (in Chinese). Chinese Science Bulletin, 43:860-862.
IGLESIAS, L.G., SOLÍS-RAMOS, L.Y. y VIVEROS-VIVEROS, H., 2012. Variación morfométrica en dos poblaciones naturales de Pinus hartwegii Lindl. del estado de Veracruz. Phyton (Buenos Aires) [en línea], vol. 81, no. 2, pp. 239-247. [Consulta: 6 mayo 2019]. ISSN 1851-5657. Disponible en: http://www.scielo.org.ar/scielo.php?script=sci_abstract&pid=S1851 -56572012000200017&lng=es&nrm=iso&tlng=pt.
JANKOWSKI, A., WYKA, T.P., ¯YTKOWIAK, R., NIHLGÅRD, B., REICH, P.B. y OLEKSYN, J., 2017. Cold adaptation drives variability in needle structure and anatomy in Pinus sylvestris L. along a 1,900 km temperateboreal transect. Functional Ecology [en línea], vol. 31, no. 12, pp. 2212-2223. [Consulta: 6 mayo 2019]. ISSN 1365-2435. DOI 10.1111/1365-2435.12946. Disponible en: https://besjournals.onlinelibrary.wiley.com/doi/abs/10.1111/1365-2435.12946.
KEELEY, J.E., 2012. Ecology and evolution of pine life histories. Annals of Forest Science [en línea], vol. 69, no. 4, pp. 445-453. [Consulta: 6 mayo 2019]. ISSN 1297-966X. DOI 10.1007/s13595-012-0201-8. Disponible en: https://doi.org/10.1007/s13595-012-0201-8.
KIVIMÄENPÄÄ, M., SUTINEN, S., VALOLAHTI, H., HÄIKIÖ, E., RIIKONEN, J., KASURINEN, A., GHIMIRE, R.P., HOLOPAINEN, J.K. y HOLOPAINEN, T., 2017. Warming and elevated ozone differently modify needle anatomy of Norway spruce Picea abies and Scots pine Pinus sylvestris. Canadian Journal of Forest Research [en línea], vol. 47, no. 4, pp. 488-499. [Consulta: 6 mayo 2019]. ISSN 0045-5067, 1208-6037. DOI 10.1139/cjfr-2016-0406. Disponible en: http://www.nrcresearchpress.com/doi/10.1139/cjfr-2016-0406.
KÖBÖLKUTI, Z.A., TÓTH, E.G., LADÁNYI, M. y HÖHN, M., 2017. Morphological and anatomical differentiation in peripheral Pinus sylvestris L. populations from the Carpathian region. . S.l.: s.n., DOI 10.12657/denbio.077.009.
LIN, J., JACH, M.E. y CEULEMANS, R., 2001. Stomatal density and needle anatomy of Scots pine (Pinus sylvestris) are affected by elevated CO2. New Phytologist [en línea], vol. 150, no. 3, pp. 665-674. [Consulta: 6 mayo 2019]. ISSN 1469-8137. DOI 10.1046/j.1469-8137.2001.00124.x. Disponible en: https://nph.onlinelibrary.wiley.com/doi/abs/10.1046/j.1469-8137.2001.00124.x.
LÓPEZ-ALMIRAL, A., 1982. Variabilidad del género Pinus (Pinaceae) en Cuba. Acta Botánica Cubana [en línea], vol. 12, no. 1, pp. 43. ISSN 2519-7754. Disponible en: http://revistas.geotech.cu/index.php/abc/index.
LUOMALA, E.M.; LAITINEN, K.; SUTINEN, S.; KELLOMÄKI, S.; VAPAAVUORI, E. 2005. Stomatal density, anatomy and nutrient concentrations of Scots pine needles are affected by elevated CO2 and temperatura. Plant, Cell and Environment, 28:733749.
MENG, J., CHEN, X., HUANG, Y., WANG, L., XING, F. y LI, Y., 2018. Environmental contribution to needle variation among natural populations of Pinus tabuliformis. Journal of Forestry Research [en línea], [Consulta: 6 mayo 2019]. ISSN 1993-0607. DOI 10.1007/s11676-018-0722-6. Disponible en: https://doi.org/10.1007/s11676-018-0722-6.
O'NEILL, G.A., AITKEN, S.N., KING, J.N. y ALFARO, R.I., 2002. Geographic variation in resin canal defenses in seedlings from the Sitka spruce × white spruce introgression zone. . S.l.: s.n., DOI 10.1139/x01-206.
REINHARDT, K., CASTANHA, C., GERMINO, M.J., KUEPPERS, L.M. y PEREIRA, J., 2011. Ecophysiological variation in two provenances of Pinus flexilis seedlings across an elevation gradient from forest to alpine. Tree Physiology [en línea], vol. 31, no. 6, pp. 615-625. [Consulta: 6 mayo 2019]. ISSN 0829-318X. DOI 10.1093/treephys/tpr055. Disponible en: https://academic.oup.com/treephys/article/31/6/615/1657213.
SATYENDRA PRAKASH, T., PRADEEP, K., DEEPIKA, Y. y DEVENDRA KUMAR, C., 2013. Comparative morphological, epidermal, and anatomical studies of Pinus roxburghii needles at different altitudes in the North-West Indian Himalayas. Turkish Journal of Botany [en línea], vol. 37, pp. 65-73. [Consulta: 6 mayo 2019]. Disponible en: https://www.academia.edu/5992716/Comparative_morphological_epidermal_and_anatomical_studies_of_Pinus_roxburghii_needles_at_different_altitudes_in_the_North -West_Indian_Himalayas.
SCHOETTLE, A.W. y ROCHELLE, S.G., 2000. Morphological variation of Pinus flexilis (Pinaceae), a bird-dispersed pine, across a range of elevations. American Journal of Botany, vol. 87, no. 12, pp. 1797-1806. ISSN 0002-9122.
SEONG, D. y OFFNER, S., 2013. A Phylogenetic Study of Conifers Describes Their Evolutionary Relationships and Reveals Potential Explanations for Current Distribution Patterns |. Journal of Emerging Investigators [en línea], pp. 1-9. [Consulta: 6 mayo 2019]. Disponible en: https://www.emerginginvestigators.org/system/articles/pdfs/000/000/053/original /6d76e2286cd825ff847f2eb1cf77ad39aa702bc3.pdf.
SHEUE, C.-R., CHANG, H.-C., YANG, Y.-P., LIU, H.-Y., CHESSON, P. y HSU, F.-H., 2014. A new classification of marginal resin ducts improves understanding of hard pine (Pinaceae) diversity in Taiwan. Flora - Morphology, Distribution, Functional Ecology of Plants [en línea], vol. 209, no. 8, pp. 414-425. [Consulta: 6 mayo 2019]. ISSN 0367-2530. DOI 10.1016/j.flora.2014.06.007. Disponible en: http://www.sciencedirect.com/science/article/pii/S0367253014000760.
ZHANG, L., NIU, H., WANG, S., ZHU, X., LUO, C., LI, Y. y ZHAO, X., 2012. Gene or environment? Species-specific control of stomatal density and length. Ecology and Evolution [en línea], vol. 2, no. 5, pp. 1065-1070. [Consulta: 6 mayo 2019]. ISSN 2045-7758. DOI 10.1002/ece3.233. Disponible en: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3399171/.
![]()
This work is licensed under a Creative Commons Attribution-NonCommercial 4.0 International license
Copyright (c) 2019 Liutmila Pérez-Del Valle, Gretel Geada
López, Rogelio
Sotolongo-Sospedra