Revista Cubana de Ciencias Forestales. May-August, 2019, 7(2): 184-196![]()
Translated from the original in spanish
Impact of mega-drought on tree radial oak growth Nothofagus obliqua (Mirb.) Oerst. (Roble), Chile
Impacto de la megasequía en el crecimiento radial de Nothofagus obliqua (Mirb.) Oerst. (Roble), Chile
Patricio Corvalán Vera1
Jaime Hernández Palma2
1Universidad de Chile. Chile. E-mail: pcorvalanvera@gmail.com
Received: February 5th, 2019.
Approved: April 3rd, 2019.
ABSTRACT
In order to evaluate the impact of the decline in rainfall between 2007 and 2012 in Central Chile on the northern forests of Nothofagus obliqua, a total of 223 growth plugs were measured in 51 isolated trees and 172 growing in competition. Local annual precipitation data were used to compare the growths of the normal period (2001-2006) with those of the dry period (2007-2012). The current annual growth in radius to normal diameter and its sensitivity were contrasted, showing that both isolated and competing trees showed an evident deterioration in their growth rates. A 34.8 % deficit in precipitation translates into a 17.6 % and 28.8 % decrease in current annual growth in isolated and competing trees, respectively. In the dry period, compared to the previous period, isolated trees showed a less sensitive growth (0.364) than that of competing trees (0.384). It was also established that the precipitation evaluated in a moving average of previous years to the year in evaluation is able to explain, through a linear model, up to 57.3 % of the total variability of the radial growth in isolated trees, using an average window of 6 years and 41.7 % in trees that compete with a window of 3 years.
Keywords: competence; tree radial growth; precipitation; drought.
RESUMEN
Con el objeto de evaluar el impacto de la declinación de las precipitaciones ocurrida entre los años 2007 al 2012 en Chile Central, sobre los bosques septentrionales de Nothofagus obliqua, se midieron un total de 223 tarugos de crecimiento en 51 árboles aislados y 172 creciendo en competencia. Se utilizaron datos locales de precipitación anual para comparar los crecimientos del período normal (2001-2006) con los del período seco (2007-2012). Se contrastó el crecimiento anual corriente en radio al diámetro normal y su sensitividad, mostrando que tanto los árboles que crecen aislados como los que compiten presentaron un evidente deterioro en sus tasas de crecimiento. Un 34.8 % de déficit en precipitación se traduce en un 17.6 % y 28.8 % de disminución en el crecimiento anual corriente en árboles aislados y en competencia, respectivamente. En el período seco, respecto al período previo, los árboles aislados presentaron un crecimiento menos sensitivo (0.364) que el de los árboles en competencia (0.384). También se estableció que la precipitación evaluada en un promedio móvil de años previos al del año en evaluación es capaz de explicar, a través de un modelo lineal, hasta el 57.3 % de la variabilidad total del crecimiento radial en los árboles aislados, usando una ventana promedio de 6 años y 41.7 % en los árboles que compiten con una ventana de tres años.
Palabras clave: competencia; crecimiento radial; precipitación; sequía.
INTRODUCTION
Climate change predictions for the Andean region of the Maule Region warn that by the end of the century there will be a 60 % to 70 % decrease in annual rainfall (Ppt) and an increase in temperature of up to 5º C, which added to the anthropic destruction generates a highly vulnerable condition for the northern montane deciduous forests dominated by N. obliqua (Oak) that grow in this area (Donoso, 1982; Lara et al., 2012).
The water deficit directly and indirectly inhibits the growth of trees (Kozlowski and Pallardi 1997, Bonan 2008, Aubry-Kientz and Moran 2017) by conditioning the energy balance between survival, vegetative and reproductive growth (Thabeet et al., 2009; Hulshof et al., 2012), which added to the impact of inter-specific competition generates a complex problem to model and quantify.
Dendrochronology is a valuable tool for measuring the impacts of the environment on tree growth and recovering information from past behavior (Fritts, 1976; Babst et al., 2013). The historical measurement of growths, together with climatic data, makes it possible to establish some basic deductions to evaluate the behavior of the species in the face of the effects of climate change and to carry out some elementary silvicultural scopes.
The prolonged deficit of precipitations occurred between the years 2007-2012 in the central zone of Chile allows to study empirically the effect it has had on the Annual Radial Current Growth (CAC) of trees, growing both in isolated form and in competition and constitutes an approximation of what could be a permanent condition in the future. The prolonged deficit of precipitations for six consecutive years constitutes an unrepeatable scenario to evaluate its impact in the growth of the forests.
The purpose of this paper is to quantify the relationships between annual precipitation, radial growth and the effect of competition, as well as to measure the sensitivity of growth rings. Another objective is to determine the length of time prior to annual growth that best explains the relationship between rainfall and current annual growth, in a normal climatic and drought period, in unmanaged secondary oak forests.
MATERIALS AND METHODS
Work area
The information was collected in the precordillerano sector of Monte Oscuro (19H 324862E-6113124S), located 50 km southeast of the city of Curicó, commune of Romeral, province of Curicó, region of Maule, Chile. The study area covers an area of approximately 1,300 hectares and corresponds to formations of secondary forests of the forest types Roble-Hualo, Ciprés de la cordillera and Roble-Raulí-Coigüe. This area was cleared for sawn timber and posts in the 1950s. Subsequently, the forest has been cleared in small areas for the production of firewood and charcoal. Hence its formation as a mosaic of secondary contemporary groves.
From a morphogenetic point of view, the soil is the product of materials from moraine deposits, fluvial and glacio-fluvial hauling, as well as some lake and glacio-lacustrian deposits that have been covered by layers of volcanic ash (CONAF, 1999). Consequently, the soil is deep, derived from volcanic ash and has a loamy texture, high organic content and low availability of phosphorus and nitrogen (CONAF, 1999).
In terms of vegetation, the area corresponds to high altitude Andean oak forests, where it is possible to find the species growing alongside Peumus boldus and Cryptocarya alba, typical of the sclerophyllous forest, as well as Laurelia sempervirens, Aextoxicon punctatum, Nothofagus glauca and Astrocedrus chilensis, among others, depending on the exposure in which it is found (Corvalán et al., 2014).
The climate is temperate Mediterranean, according to the climatic classification of Köppen with dry season of four to five months. It is characterized by concentrated rainfall during the winter months, while in the months of the dry season (October to March) summer rainfall does not exceed 40 mm per month. The foothills have occasional snowfalls in winter (Atmosphere, 2014).
In terms of land cover, a LiDAR data set with a density of 4.64 points per m2 (p*m-2) and a VIS aerial image with a spatial resolution of 1 m were acquired and orthorectified in March 2011. From this information, a detailed digital elevation model of terrain and cups was available that allowed the measurement of the model of heights with very high spatial resolution (1 m). There are also aerial metric photographs of the Chilean Aerophotogrammetric Service of 1980 and 1995-1996, scales 1:50 000 and 1:20 000, respectively, and color photos of 1991, scale 1:5 000 nonmetric. Two intensive forest inventories were also carried out between 2011 and 2013.
Measurement of radial growth rings
The dendrometric samples that were collected correspond to 51 trees that grow in isolation and 172 trees that grow in full competition. The samples were taken under different conditions of size and stage of development of the stands. Spatial selection was made using the terrain elevation and ground cover model and aerial imagery and forest inventories (Figure 1). The competing trees come from 43 forest inventory plots. In each plot, the species was identified and normal diameters (NAD) were measured for trees larger than 10 cm DBH. Once measured they were ordered in increasing values of DBH and from each quartile of the population the central individual was selected to which a growth plug was extracted in direction to the center of the plot with drill mark JIM-GEM.
The dowels were dried in the open air, placed on a wooden base and sanded with three different grains to facilitate their measurement. The readings were made with 400X CoolingTech electron microscope. Current annual growth was measured in radius (mm) between 2001 and 2012 at DAP level. This resulted in a matrix of 223 rows (51 isolated trees plus 172 competing trees) per 12 columns (years 2001 to 2012). CCS data at tree level were averaged annually.

Fig. 1.- Map with location of trees in the study area
In red, the trees growing in competition and in yellow isolated trees
Annual rainfall
Monthly precipitation data from the meteorological station "Monte Oscuro", belonging to the Dirección General de Aguas del Ministerio de Obras Públicas del Gobierno de Chile, located about 3.5 km from the study area in the same basin, downstream, between 2001 and 2012 (DGA 2014), were used. Annual rainfall data were accumulated monthly.
Autocorrelations and sensitivity
To measure the level of self-reliance of the CLC in DAP, the matrix of historical correlations of average CLCs for isolated trees and those growing in competition was analysed (Figure 2). Sensitivity was calculated as:
S(i+1)=(ABS(CAC(i+1) - CACi)*2)/(CAC(i+1) + CACi) [1]
Where:
S(i+1) is the sensitivity of the current annual growth between years i and (i+1) (Schweingruber 1988).
Relationships current annual growth vs. precipitation previous years
Current annual growth data were averaged over the years 2001-2006 (normal period) and compared with the period 2007-2012 (mega-droughts). The current annual growth was related to the moving average of precipitations occurred in "j" previous years (between 1994 and 2012). A linear model was adjusted to find the number of years that would maximize the variation in growth explained by the regression (R2). The model was as follows (Equation 1).
CACi=a+b*Ppt (j) [2].
Where:
a, b adjustment parameters,
CACi is the average current annual growth (mm*year -1) in radius in the year
i, Ppt(j) is the average annual rainfall in the last j years (mm* year -1)
RESULTS AND DISCUSSION
The current annual growth in radius for the study period is presented in Table 1 (Table 1).
Table 1.- Precipitation, current annual growth and sensitivity of the analysis period

From table 1 it can be deduced that a drop in precipitation from 1 630 mm*year-1 to 1 062 mm* year-1 reduces the average growth rate from 3,811 mm* year-1 to 3,241 mm* year-1 in isolated growing trees (17.6 %) and from 1,812 mm* year-1 to 1,407 mm* year-1 in competing growing trees (28.8 %).
According to Donoso et al., (2010), the radial growth of a marginal Nothofagus macrocarpa forest located in central Chile is 1.15 mm* year-1, which is much lower than the value found in this study, both in isolated and competing trees. Donoso (1988), studying oak renewals in an area close to those of this study, determined annual radial growths for 10-year periods between 1.75 and 3.45 mm* year-1 , values higher than those found in this study in competing trees. In a more southern area, De la Maza and Gilchrist (1983) found radial growth values between 2.5 and 4 mm* year -1, higher than those found in this study. The results reported are consistent with the precipitation gradient along the latitudinal distribution in Chile. No similar study of the same species has been reported in the literature to compare the impact of rainfall decline, even when comparing the average CAC data of this study with those reported by Adams and Kolb (2005), who, by studying the ratio of growth in wet versus dry years in Pinus ponderosa, Pseudotsuga menziesii, Pinus flexilis and Picea engelmannii, in northern Arizona reported similar values in all sites, species and communities with that of this study (Adams and Kolb, 2005).
Table #1 shows the average sensitivity values, showing a decrease in sensitivity in isolated trees during drought and, conversely, an increase in sensitivity in trees growing in competition. The decrease in sensitivity of isolated trees to drought would be explained by a gradual decrease in water reserves in the deep soil, making the annual growth more homogeneous under these conditions. On the other hand, the increase in the sensitivity of growth in competing trees is explained by the effect of drought between the different social conditions in which individuals grow. The difference in growth between individuals of different percentiles (of DAP) is accentuated in dry periods, making the driest years more sensitive. (2004), who studying Nothofagus dombeyi (coihue) in the Nahuel Huapi National Park in northeastern Patagonia, in a year of extreme drought, found that juvenile individuals presented growth patterns more sensitive and prone to mortality than older trees (Suárez et al., 2004).
Figure #2 shows the variation in annual rainfall and CCS trends for both isolated and competing trees. It is observed that there is a delay effect of the impact of rainfall on the growth pattern of trees, where isolated trees clearly grow more because they do not have competition effect. The precipitations on the sensitivity pattern with a delay effect of between 1 and 2 years of growth trends in both cases are similar. A lag effect is also observed (Figure 2).
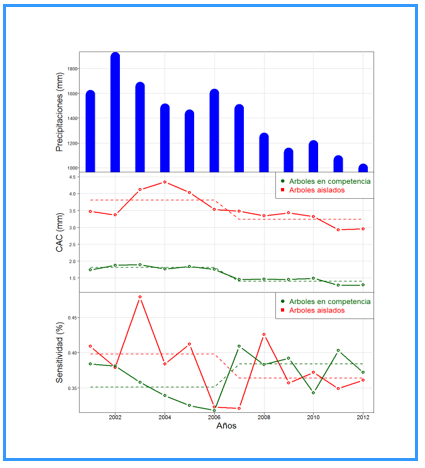
Fig. 2. - Annual rainfall (mm), current annual growth in radius (mm/year)
and annual sensitivity (%) of the CAC in radius for isolated and competing trees.
The horizontal segmented lines are the values of the annual average for the normal period (2001-2006) and dry period (2007-2012)

Fig. 3. - Matrix of CAC correlations in radius between different years
for the normal rainfall period (2001-2006) and for the dry period (2007-2012)
Above: Isolated growing trees. Below: trees growing in competition.
The values above the diagonal plot the Spearman correlation coefficient in color ranges.
Absolute correlation values are indicated below the diagonal.
The years in which the correlation was established are indicated on the diagonal. All of them are significant with an error alpha=0.05
One of the most notable effects of precipitation on tree growth is shown in Figure 3, which shows the close autocorrelation of growths in the period studied. Trees growing in competition show a higher level of interdependence between successive growths than trees growing in isolation. This phenomenon is well described by Fritz (1976), who points out that in trees there is a dynamic energy balance in vegetative growth. The precipitations of one year condition the growth of later years (Fritts, 1976; Kozlowski and Pallardi 1997). Water deficit directly and indirectly inhibits tree growth by conditioning the energy balance between survival, vegetative and reproductive growth (Thabeet et al., 2009; Hulshof et al., 2012) (Figure 3).
Table #2 shows the fit parameters of the [2] model of current annual growth vs. average precipitation in "j" years (Table 2).
Table. 2 - Current annual growth vs. average precipitation in "j" previous years

It can be observed that, under this simple linear model, average rainfall decreases adjustment errors by incorporating more years, explaining 57.3 % of the total variance of error in the case of isolated trees, using a six-year mobile sale and 41.7 % in the case of competing trees using three years. This information is of interest if it is related to the autocorrelation matrix indicated in figure 3. In both cases, it can be interpreted as a positive effect of water storage in the soil. Some studies that relate growth to precipitation have been especially developed in tropical areas. Brienen and Zuidema (2005) find a determination coefficient of 7.9 % for Cedrelinga catenaeformis, a legume that grows in a rainforest in Bolivia, when estimating the width of the standardized growth ring as a function of total precipitation. (1995) find in Central Panama determination coefficients between 28 % and 44 % for Pseudobombax septenatum, Cordia alliodora and Annona spraguei. There is no information available to compare growth with the cumulative time effect of soil water storage.
The 34.8 % decrease in average annual precipitation (2006-2012) (from 1 630 to 1 062 mm* year -1 ) for the period (2001-2006) generates a fall of:
The average annual precipitation as a predictor variable of the CAC, is able to explain that:
The autocorrelations of the CAC are significant (at 95 % reliability) and reach the whole analysis period (12 years), both for isolated trees and for competing trees, covering both normal years of precipitation and dry years.
In the dry period, compared with the normal period:
BIBLIOGRAPHICAL REFERENCES
ADAMS, H.D. y KOLB, T.E., 2005. Tree growth response to drought and temperature in a mountain landscape in northern Arizona, USA. Journal of Biogeography [en línea], vol. 32, no. 9, pp. 1629-1640. [Consulta: 27 febrero 2019]. ISSN 0305-0270. DOI 10.1111/j.1365-2699.2005.01292.x. Disponible en: https://nau.pure.elsevier.com/en/publications/tree-growth-response-to-drought-and-temperature-in-a-mountain-lan.
AUBRY-KIENTZ, M. y MORAN, E.V., 2017. Climate Impacts on Tree Growth in the Sierra Nevada. Forests [en línea], vol. 8, no. 11, pp. 414. [Consulta: 27 febrero 2019]. DOI 10.3390/f8110414. Disponible en: https://www.mdpi.com/1999-4907/8/11/414.
Atmósfera., 2014 Información climatológica de estaciones chilenas. Departamento de Geofísica de la Facultad de Ciencias Físicas y Matemáticas de la Universidad de Chile. Disponible en: http://www.atmosfera.cl/HTML/datos/datos_02.html
BABST, F., POULTER, B., TROUET, V., TAN, K., NEUWIRTH, B., WILSON, R., CARRER, M., GRABNER, M., TEGEL, W., LEVANIC, T., PANAYOTOV, M., URBINATI, C., BOURIAUD, O., CIAIS, P. y FRANK, D., 2013. Site- and species-specific responses of forest growth to climate across the European continent. Global Ecology and Biogeography [en línea], vol. 22, no. 6, pp. 706-717. [Consulta: 9 mayo 2019]. ISSN 1466-8238. DOI 10.1111/geb.12023. Disponible en: https://onlinelibrary.wiley.com/doi/abs/10.1111/geb.12023.
BONAN, G.B., 2008. Forests and Climate Change: Forcings, Feedbacks, and the Climate Benefits of Forests. Science [en línea], vol. 320, no. 5882, pp. 1444-1449. [Consulta: 27 febrero 2019]. ISSN 0036-8075, 1095-9203. DOI 10.1126/science.1155121. Disponible en: http://science.sciencemag.org/content/320/5882/1444.
BRIENEN, R.J.W. y ZUIDEMA, P.A., 2005. Relating tree growth to rainfall in Bolivian rain forests: a test for six species using tree ring analysis. Oecologia, vol. 146, pp. 1-12. DOI 10.1007/s00442-005-0160-y.
Corporación Nacional Forestal (CONAF). 1999. Documento de trabajo N° 298 plan de manejo Reserva Nacional Los Bellotos del Melado. Unidad de Gestión Patrimonio Silvestre. 71p.
CORVALÁN, P., GALLEGUILLOS, M. y HERNÁNDEZ, J., 2014. Presencia, abundancia y asociatividad de Citronella mucronata en bosques secundarios de Nothofagus obliqua en la precordillera de Curicó, región del Maule, Chile. Bosque (Valdivia) [en línea], vol. 35, no. 3, pp. 269-278. [Consulta: 27 febrero 2019]. ISSN 0717-9200. DOI 10.4067/S0717-92002014000300002. Disponible en: https://scielo.conicyt.cl/scielo.php?script=sci_abstract&pid=S0717-92002014000300002&lng=en&nrm=iso&tlng=es.
DE LA MAZA, C. y GILCHRIST, J., 1983. Algunos antecedentes para el manejo de renovales de Raulí. Santiago. Facultad de Ciencias Forestales de la Universidad de Chile, vol. Boletín Técnico, (61), no. 30.
DEVALL, M.S., PARRESOL, B.R. y WRIGHT, S.J., 1995. Dendroecological Analysis of Cordia Alliodora, Pseudobombax Septenatum and Annona Spraguei in Central Panama. IAWA Journal [en línea], vol. 16, no. 4, pp. 411-424. [Consulta: 27 febrero 2019]. ISSN 0928-1541, 2294-1932. DOI 10.1163/22941932-90001430. Disponible en: https://brill.com/abstract/journals/iawa/16/4/article-p411_10.xml.
DIRECCIÓN GENERAL DE AGUAS (DGA), 2014. Información Oficial Hidrometeorológica y de Calidad de Aguas en Línea [en línea]. 2014. S.l.: DIRECCIÓN GENERAL DE AGUAS (DGA). [Consulta: 15 mayo 2014]. Disponible en: http://snia.dga.cl/BNAConsultas/.
DONOSO Z, C., 1982. RESEÑA ECOLOGICA DE LOS BOSQUES MEDITERRANEOS DE CHILE. Bosque (Valdivia) [en línea], vol. 4, no. 2, pp. 117-146. [Consulta: 27 febrero 2019]. ISSN 0717-9200. Disponible en: http://mingaonline.uach.cl/scielo.php?pid=S0717-92001982000100004&script=sci_arttext.
DONOSO, P., 1988. Caracterización y proposiciones silviculturales para renovales de Roble (Nothofagus obliqua) y Raulí (Nothofagus alpina) en el área de protección "RadalSiete Tazas". Bosque, 9(2), 103-114. Disponible en: https://bibliotecadigital.infor.cl///handle/20.500.12220/8855
DONOSO, S.R., PEÑA-ROJAS, K., DELGADO-FLORES, C., RIQUELME, A. y PARATORI, M., 2010. ABOVE-GROUND BIOMASS ACCUMULATION AND GROWTH IN A MARGINAL Nothofagus macrocarpa FOREST IN CENTRAL CHILE. Interciencia [en línea], vol. 35, no. 1, pp. 65-69. [Consulta: 27 febrero 2019]. ISSN 0378-1844, 2244-7776. Disponible en: http://www.redalyc.org/articulo.oa?id=33913146011.
FRITTS, H., 1976. Tree Rings and Climate [en línea]. 1st Edition. S.l.: Academic Press. [Consulta: 27 febrero 2019]. ISBN 978-0-323-14528-2. Disponible en: https://www.elsevier.com/books/tree-rings-and-climate/fritts/978-0-12-268450-0.
HULSHOF, C.M., STEGEN, J.C., SWENSON, N.G., ENQUIST, C.A.F. y ENQUIST, B.J., 2012. Interannual variability of growth and reproduction in Bursera simaruba: the role of allometry and resource variability. Ecology, vol. 93, no. 1, pp. 180-190. ISSN 0012-9658. Disponible en: https://www.ncbi.nlm.nih.gov/pubmed/22486098
KOZLOWSKI, T.T. y PALLARDY, S.G., 1997. Growth control in woody plants [en línea]. San Diego: Academic Press. [Consulta: 27 febrero 2019]. ISBN 978-0-12-424210-4. Disponible en: http://site.ebrary.com/id/10187296.
Lara, A., Solari, M.E., Prieto, M.D.R. y Peña, M.P., 2012. Reconstrucción de la cobertura de la vegetación y uso del suelo hacia 1550 y sus cambios a 2007 en la ecorregión de los bosques valdivianos lluviosos de Chile (35º-43º 30´S). Bosque (Valdivia), 33(1): 13-23.
SCHWEINGRUBER, F.H., 1988. Tree Rings: Basics and Applications of Dendrochronology [en línea]. S.l.: Springer Netherlands. [Consulta: 9 mayo 2019]. ISBN 978-0-7923-0559-0. Disponible en: https://www.springer.com/us/book/9780792305590.
SUAREZ, M.L., GHERMANDI, L. y KITZBERGER, T., 2004. Factors predisposing episodic drought-induced tree mortality in Nothofagus site, climatic sensitivity and growth trends. Journal of Ecology [en línea], vol. 92, no. 6, pp. 954-966. [Consulta: 9 mayo 2019]. ISSN 1365-2745. DOI 10.1111/j.1365-2745.2004.00941.x. Disponible en: https://besjournals.onlinelibrary.wiley.com/doi/abs/10.1111/j.1365-2745.2004.00941.x.
Thabeet, A., Vennetier, M., Gadbin-Henry, C., Denelle, N., Roux, M., Caraglio, Y. y Vila, B., 2009. Response of Pinus sylvestris L. to recent climatic events in the French Mediterranean Region. Trees, 23(4), 843-853. Disponible en: https://doi.org/10.1007/s00468-009-0326-z
![]()
This work is licensed under a Creative Commons Attribution-NonCommercial 4.0 International license
Copyright (c) 2019 Patricio Corvalán Vera, Jaime Hernández
Palma