Revista Cubana de Ciencias Forestales. 2018; enero-abril, 6(1):15-30
![]()
Proyección de supervivencia en plantaciones de Pinus caribaea Morelet var. caribaea Barrett & Golfari
Survival prognosis in plantations of Pinus caribaea Morelet var. caribaea Barrett & Golfari
Ouorou Ganni Mariel Guera1
José Antônio Aleixo da Silva1
Rinaldo Luiz Caraciolo Ferreira1
Daniel Álvarez Lazo2
Héctor Barrero Medel2
1Universidad Federal Rural de Pernambuco (UFRPE), Departamento de Ciência
Florestal (DCFL), Laboratório de Biometria e Manejo Florestal (LBMF). Brasil.
Correo
electrónico: gueraforest@gmail.com; jaaleixo@gmail.com; rinaldo.ferreira@ufrpe.br
2Ingeniero Forestal. Universidad de Pinar del Río «Hermanos Saíz Montes de Oca»,
Facultad de Ciencias Forestales y Agropecuaria Departamento Forestal. Cuba.
Correo
electrónico: daniel@upr.edu.cu; hbarrero@upr.edu.cu
Recibido: 2 de noviembre 2017.
Aprobado: 30 de enero 2018.
RESUMEN
El presente estudio fue realizado con el objetivo de obtener ecuaciones de regresión y Redes Neuronales Artificiales (RNAs) para la proyección de la supervivencia de Pinus caribaea var. caribaea en la empresa forestal Macurije, provincia de Pinar del Río, Cuba. Los datos utilizados en las modelaciones provienen de la medición de las variables edad (años) y supervivencia (densidad) en parcelas permanentes circulares de 500 m², establecidas en plantaciones de P. caribaea var. caribaea. El estudio se dividió en tres etapas: 1) Ajuste de modelos de regresión tradicionales para proyección de supervivencia; 2) Entrenamientos de RNAs para la proyección de supervivencia, incluyendo variables categóricas «sitio» y «Unidades Básicas de Producción Forestal»; 3) Comparación de los desempeños de las ecuaciones de regresión con los de las RNAs en la proyección de la supervivencia. Los mejores modelos y RNAs fueron seleccionados y basados en: coeficiente de determinación ajustado -R2aj (%), raíz cuadrada del error medio cuadrático - RMSE (%) y análisis de distribución de residuos. La evaluación de la bondad de ajuste de los modelos también incluyó la verificación de los supuestos de normalidad, homocedasticidad y ausencia de autocorrelación serial en los residuos por las pruebas de Kolmogorov-Smirnov, White y Durbin-Watson, respectivamente. El modelo de Pienaar y Shiver de 1981 resultó ser el de mejor ajuste en la proyección de la supervivencia. La RNA de arquitectura MLP 13-10-1 fue la de mejor capacidad de generalización y presentó un desempeño similar al de la ecuación obtenida del ajuste del modelo de Pienaar y Shiver.
Palabras clave: Plantaciones forestales; mortalidad regular; regresión no lineal; Redes Neuronales Artificiales.
ABSTRACT
The present study was carried out with the objective of obtaining regression equations and Artificial Neural Networks (ANNs) for the prognosis of Pinus caribaea var. caribaea survival in Macurije Forest Company, province of Pinar del Río - Cuba. The data used in the modeling comes from the measurement of the variables age (years) and survival (density) in circular permanent plots of 500 m² established in P. caribaea var. caribaea plantations. The study was divided into three stages: i) Adjustment of survival traditional regression models; ii) Training of ANNs for survival prognosis, including categorical variables «site» and «Basic Units of Forest Production»; iii) Comparison of regression equations performance with those of ANNs in survival prognosis. The best models and ANNs were selected based on: adjusted determination coefficient - R2aj (%), square root of the mean square error - RMSE (%) and residue distribution analysis. The evaluation of the models goodness of fit also included the verification of the assumptions of normality, homocedasticity and absence of serial autocorrelation in the residues by Kolmogorov-Smirnov, White and Durbin-Watson tests, respectively. The model of Pienaar and Shiver 1981 turned out to be the best fit in survival prognosis. The ANN MLP 13-10-1 was the one with the best generalization capacity and presented a performance similar to that of Pienaar and Shiver equation.
Keywords: Forest plantations; regular mortality; nonlinear regression; Artificial Neural Networks (ANNs).
INTRODUCCIÓN
La mortalidad de árboles es uno de los principales elementos de la dinámica forestal Bigler, (2003). La misma, como variable complementaria de la supervivencia, puede ser definida como la reducción de la densidad de un bosque o de una plantación, ocasionada por factores naturales Sanquetta, (1990). Según Campos y Leite (2017), este fenómeno (la mortalidad) no debe confundirse con las fallas de siembra que ocurren en la etapa inicial del establecimiento de las plantaciones y que pueden ser repuestas. Descuidar la mortalidad, como sucede generalmente en los estudios de crecimiento y producción forestal, es aceptable solo en plantaciones forestales industriales intensamente manejadas Vanclay, (1991), principalmente, si los raleos periódicos están incluidos dentro de los tratamientos forestales aplicados Campos y Leite, (2017).
Aunque este componente de crecimiento sea implícito en los modelos de proyección de crecimiento y producción forestal, su modelación se realiza de forma explícita para obtener ecuaciones de auxilio a la toma de decisiones relacionadas con las densidades de los rodales. En dependencia de la fuente de los factores naturales al que hace referencia Sanquetta (1990), Lee (1971) distingue dos tipos de mortalidad: la mortalidad regular, característica de plantaciones muy maduras y densas, es atribuida a factores intrínsecos, a los árboles como la senescencia, la genética y la alta competencia por recursos naturales limitados; y la mortalidad irregular, causada por factores no intrínsecos al árbol como incendios, plagas y enfermedades, vientos y daños mecánicos. La complejidad de la previsión de los factores causantes de la mortalidad irregular dificulta su proyección, por lo que la mayoría de los estudios se enfocan en la mortalidad regular, interés del presente estudio.
Numerosos métodos de proyección de mortalidad regular se desarrollaron para los bosques monoespecíficos (plantaciones puras) Vanclay, (1991). Los mismos son generalmente propuestos al nivel de plantación o de árbol individual Thapa, (2014). Los modelos al nivel de plantación se basan en la edad y densidad inicial del rodal y los desarrollados al nivel de árbol individual se basan, generalmente, en la probabilidad de mortalidad de cada árbol o grupo de árboles. Las variables dependientes de estos modelos son: el número de árboles por hectárea (N/ha.), como en los modelos desarrollados por Clutter y Jones (1980), Pienaar y Shiver (1981) o Silva y Bailey (1986) que proyectan la supervivencia (variable complementaria de la mortalidad) al nivel de plantación y la probabilidad de mortalidad de los árboles como en el enfoque propuesto por Hamilton (1974) al nivel de árboles individuales y la tasa anual de supervivencia por clase diamétrica como en el enfoque de Buchman et al. (1983) para grupos de árboles.
Entre los modelos, generalmente no lineales, propuestos para la modelación de la supervivencia (o mortalidad) al nivel de plantación, se pueden destacar los de Clutter y Jones (1980), Pienaar y Shiver (1981) y Silva y Bailey (1986) que se han mostrado adecuados y eficientes para el análisis del fenómeno en numerosos estudios. Según Clutter et al. (1983), todo modelo adecuado para la proyección de mortalidad debe tener las siguientes propiedades: (1) Si la edad final (I2) es igual a la edad inicial (I1), la densidad o sobrevivencia final (N2) debe ser igual a la densidad o sobrevivencia inicial (N1); (2) Para rodales coetáneos, cuando I2 es muy avanzada, N2 debe aproximarse a cero; (3) Si el modelo se utiliza para proyectar N2 en la edad I2, y I2 y N2 se utilizan para proyectar N3 a una edad I3 (I3> I2> I1), el resultado debe ser igual a la proyección directa de I1 a I3.
La importancia de la proyección precisa de la mortalidad en los sistemas de crecimiento y producción forestal es irrefutable; pero la mortalidad sigue siendo uno de los componentes menos comprendidos debido a la complejidad de los ecosistemas forestales Hamilton, (1986) y, debido a lo mismo, es el componente de crecimiento forestal más difícil y menos confiable de estimar Glover y Holl, (1979).
En las últimas décadas, buscando estimaciones más precisas, técnicas como las de Redes Neuronales Artificiales (RNAs) fueron ampliamente utilizadas en el sector forestal, con resultados satisfactorios. Según Haykin (2007), las redes neuronales artificiales son procesadores constituidos por unidades de procesamiento simple, que tienen la propensión natural para almacenar conocimiento experimental y hacer que esté disponible para su uso. Con el fin de una mejor comprensión de la mortalidad de los árboles y el desarrollo de modelos con mejores capacidades predictivas, numerosas investigaciones utilizaron las RNAs para la modelación de la mortalidad o supervivencia de los árboles Guan y Gertner, (1991), King et al., (2000).
Teniendo en cuenta lo anterior, el presente estudio tuvo como objetivo: ajustar modelos de regresión no lineal y entrenar RNAs para la proyección de la supervivencia en plantaciones de Pinus caribaea Morelet var. caribaea Barr. & Golf., en la Empresa Forestal Integral (EFI) Macurije, Pinar del Río, Cuba.
MATERIALES Y MÉTODOS
Ubicación del área de estudio
La presente investigación fue desarrollada en plantaciones de Pinus caribaea var. caribaea de la Empresa Forestal Integral (EFI) Macurije, ubicada aproximadamente entre las coordenadas geográficas 22º 06' a 22º 42' latitud norte y 83º 48' a 84º 23' longitud oeste, en la región más occidental de la provincia de Pinar del Río, Cuba (Fgura 1).
Fuentes de datos para la modelación de la supervivencia
Los datos fueron obtenidos de 14 parcelas permanentes de 500 m² (r = 12,615 m.), distribuidas en las dos unidades silvícolas (Guane y Mantua) y cinco unidades básicas de producción forestal (Los Ocujes, Las Cañas, Sábalo, Río Mantua y Macurije) de la Empresa Forestal Integral (EFI) Macurije. Las mismas cuentan con mediciones de las variables diámetro a altura de pecho (cm.), altura total (m.) y supervivencia (número de árboles/ha.) en diferentes edades (I). Los modelos de supervivencia se basaron y utilizaron, como variable dependiente, la densidad futura (N2) y como variables independientes la densidad inicial (N1) y las edades presentes (I1) y futuras (I2).
Modelos de proyección de supervivencia en plantaciones de P. caribaea var. caribaea en la EFI Macurije
La literatura forestal cuenta con varios modelos desarrollados para la proyección de la supervivencia/mortalidad en plantaciones forestales. Entre aquellos, que presentaron buenos ajustes a lo largo de numerosos estudios empíricos, fueron seleccionados y ajustados los de la tabla 1 (Tabla 1).

El ajuste de los modelos, así como el entrenamiento de las RNAs, abordado en la próxima sección, se realizó con el software STATISTICA vers. 8. Los modelos de supervivencia (Tabla 2) fueron ajustados por medio de los métodos iterativos de Levenberg-Marquardt, Gauss-Newton o Newton-Raphson en dependencia de la convergencia de los mismos.
Entrenamiento de Redes Neuronales Artificiales para la proyección de la supervivencia en plantaciones de Pinus caribaea var. caribaea
Se procedió al entrenamiento de 100 Redes Neuronales Artificiales (RNAs) de los tipos Multi-Layer Perceptron (MLP) y Radial Basis Function (RBF) para la proyección de supervivencia a nivel de plantación, y las mejores RNAs fueron retenidas para análisis. La supervivencia futura (N2) fue proyectada a partir de la densidad actual (N1) de las edades presentes (I1) y futuras (I2) y del índice de sitio (S). El algoritmo de entrenamiento utilizado fue el Broyden-Fletcher-Goldfarb-Shanno (BFGS), conocido por su mayor tasa de convergencia y capacidad de búsqueda más inteligente. Durante los entrenamientos, las funciones de activación probadas en las neuronas ocultas o de salida fueron las funciones Identidad (Expresión 01), Logística (Expresión 02), Exponencial (Expresión 03) y Tangente hiperbólica (Expresión 04) para las RNAs del tipo MLP.

El uso de variables cualitativas o categóricas es una de las grandes ventajas de las Redes Neuronales Artificiales. Por ello, dos variables dummies fueron incluidas en los entrenamientos: la variable «índice de sitio» para la realización de las predicciones por clase de sitio y la variable «UBPF» para posibilitar la realización de las predicciones de supervivencia por Unidad Básica de Producción Forestal (UBPF). Las categorías de esta última variable son las cinco UBPFs de la Empresa: Los Ocujes, Las Cañas, Sábalo, Río Mantua y Macurije. Los datos de las clases de sitios fueron obtenidos, a partir de las curvas polimórficas de índice de sitio, propuestas para el área de estudio. Ambas variables categóricas fueron codificadas y normalizadas por medio del procedimiento 1-de-N.
Para los entrenamientos, la base de datos fue dividida, siguiendo estas proporciones: 50 % para el entrenamiento, 25 % para la evaluación del desempeño de la red y 25 % para la validación cruzada, criterio de parada del entrenamiento de la RNA. Para mejorar la eficiencia (convergencia estable de los pesos y desvíos) del algoritmo de entrenamiento que puede verse afectado por la variación de las escalas de las variables, los datos fueron normalizados y escalados en los intervalos [0,1] o [-1,1], en dependencia de la función de activación, utilizando una transformación lineal (Expresión 07).

Para un mejor entendimiento de la relación entre las variables de entradas de las RNAs
y, principalmente, su importancia en la predicción de la supervivencia, fue realizado
un análisis sensibilidad. La contribución porcentual de
cada variable ![]() fue calculada con las expresiones 08
y 09.
fue calculada con las expresiones 08
y 09.

Criterios de evaluación y selección de los modelos
Los desempeños de los modelos de regresión y de las RNAs fueron evaluados sobre la base de coeficiente de determinación ajustado - (Expresión 10), raíz cuadrada del error medio cuadrático - RMSE (%) (Expresión 11), Sesgo (Expresión 13) y análisis de distribución de residuos (Expresión 12) para análisis de posibles tendencias de estimación (sesgos) en las ecuaciones o RNAs obtenidas.
La evaluación de los modelos de regresión fue basada, también, en la satisfacción de los supuestos de normalidad, homocedasticidad y ausencia de autocorrelación serial en los residuos verificados por las pruebas de Kolmogorov-Smirnov, White y Durbin-Watson, respectivamente.

La comparación del desempeño del modelo de mejor ajuste con el de la RNA seleccionada fue realizada con la prueba t para datos pareados.
RESULTADOS Y DISCUSIÓN
Modelos de proyección de supervivencia futura en plantaciones de Pinus caribaea
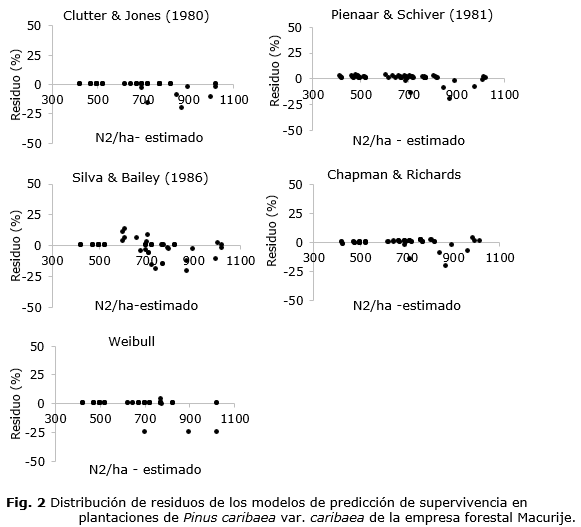
Los resultados de los ajustes de los modelos de supervivencia (tabla 2) indicaron buenos ajustes con todos los R² mayores que 97 % y RMSE menores que 5 %. Sin embargo, los valores de los Sesgos y los gráficos de distribución de residuos (figura 2) indicaron que las proyecciones menos sesgadas fueron obtenidas con la ecuación de Pienaar y Shiver (1981). En coherencia con Silva (2015), los modelos de Silva y Bailey (1986) y Chapman-Richards Chapman, (1961); Richards, (1959) presentaron desempeños similares.
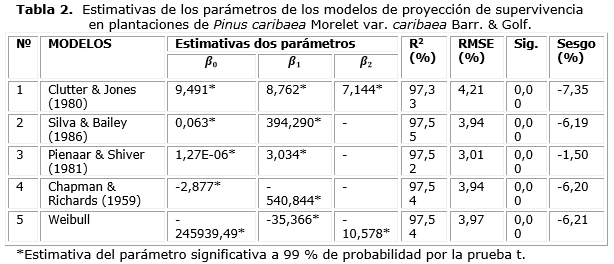
Los resultados de las pruebas de Kolmogorov-Smirnov indicaron que solo los residuos de los modelos de Silva y Bailey (1986) y Pienaar y Shiver (1981) siguieron una distribución normal, con p-valores de 0,053 y 0,068, respectivamente. En cuanto a la prueba de Durbin-Watson, sus resultados indicaron que los modelos que presentaron residuos libres de autocorrelación serial fueron los de Clutter y Jones (1980), Silva y Bailey (1986) y Pienaar y Shiver (1981) con la estadística DW igual a 1,700, 1,762 e 1,718, respectivamente. Los modelos de Chapman-Richards y de Weibull, además de presentar residuos con autocorrelación serial positiva, fueron los únicos que no atendieron el supuesto de homocedasticidad.
La satisfacción de todos los supuestos en el modelo preseleccionado (Pienaar y Shiver) favoreció su selección definitiva para la proyección de supervivencia en las plantaciones de Pinus caribaea var. caribaea de la empresa forestal Macurije.
Las proyecciones de supervivencia/mortalidad para diferentes densidades iniciales en las plantaciones de Pinus caribaea var. caribaea por medio de la ecuación de Pienaar y Schiver generaron resultados consistentes (Figura 3). Se observaron, para mayores densidades iniciales (N1), una mayor inclinación de la curva, consecuencia de una mayor tasa de mortalidad.
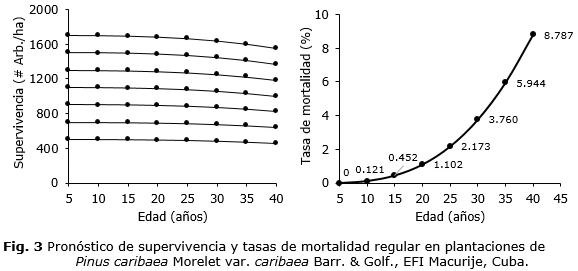
Es perceptible en la figura 3, como es de esperarse, un aumento de la tasa de mortalidad regular con el envejecimiento de las plantaciones. Las tasas de mortalidad variaron entre el 0,12 % a los 10 años a 8,78 % a los 40 años, siguiendo una forma J (o U), característica del fenómeno (mortalidad regular) estudiado Lorimer et al., (2001); Maleki y Kiviste, (2016).
Proyección de la supervivencia del Pinus caribaea var. caribaea con RNAs
La utilización de las RNAs para la predicción de la supervivencia en las plantaciones de Pinus caribaea var. caribaea proporcionó resultados satisfactorios (Tabla 3). La RNA1 de arquitectura MLP 13-10-1 presentó el mejor desempeño con proyecciones precisas y no tendenciosas (Figura 4).

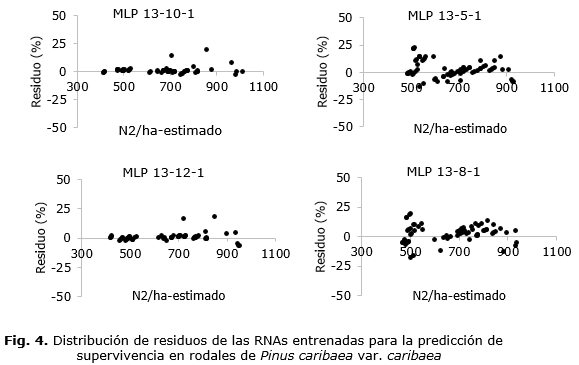
En cuanto a la importancia de las variables en los entrenamientos, el análisis de sensibilidad indicó que la variable de mayor importancia en la proyección de la supervivencia (N2) fue la densidad inicial (N1) con una importancia de 0,818. Esto explica la razón por la cual es la principal variable utilizada en la predicción de la supervivencia. Las edades presentes (I1) y futuras (I2), también ampliamente utilizadas, presentaron importancias de 0,058 y 0,045, respectivamente. De las dos variables categóricas utilizadas, la más importante fue la UBPF con un peso de 0,051.
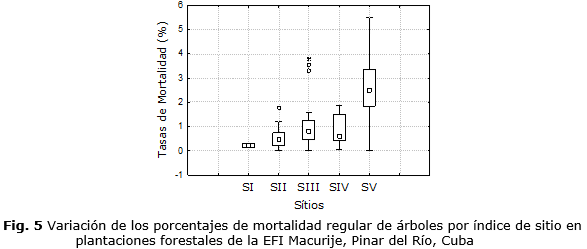
La variable menos contribuyente en la proyección de la supervivencia fue el índice de sitio con una importancia de 0,028. Hasta la fecha, el efecto del índice de sitio sobre la mortalidad de los árboles no está claro. La alta variabilidad de las tasas de mortalidad observada en algunos sitios (figura 5), principalmente los sitios de bajas capacidades productivas, indica que la calidad del sitio, por sí sola, no es suficiente para explicar la mortalidad regular o emitir consideraciones conclusivas al respecto. Cualquier tendencia observada será el resultado de la combinación de los diversos factores que afectan el crecimiento y la supervivencia de los árboles Campos y Leite, (2017). Entre esos factores, están la disponibilidad de luz, agua y nutrientes que están relacionados con la calidad de sitio y son determinantes de la mortalidad regular. A esos factores, se pueden sumar los incendios, las tempestades, los vientos, las epidemias y los ataques de insectos que son causas de la mortalidad irregular, también conocida como mortalidad catastrófica de acuerdo con la clasificación de Hann (1980).
Numerosas investigaciones empíricas llegaron a resultados divergentes que se centran en tres ejes principales: (1) la calidad del sitio influye poco en la mortalidad de los árboles; (2) la mortalidad es directamente proporcional a la calidad del sitio Vanclay, (1994), es decir, que en los mejores sitios se registran mayor mortalidad como consecuencia de una mayor competencia; y (3) la mortalidad es inversamente proporcional a la calidad del sitio, es decir, mayor mortalidad en peores sitios por poca disponibilidad de recursos.
La prueba t-pareada no fue significativa en ninguna de las comparaciones (Tabla 4), lo que indica que ambos modelos fueron eficientes en la proyección de la supervivencia. Sin embargo, los valores de los criterios de evaluación, principalmente los de RMSE (Tablas 3 y 7), indicaron una mayor precisión de la red neuronal MLP 13-10-1.

La ecuación resultante del ajuste del modelo de Pienaar y Shiver (1981) fue la más adecuada para la predicción de la supervivencia en las plantaciones de Pinus caribaea var. caribaea y presentó un desempeño similar al de la RNA MLP 13-10-1 seleccionada.
La flexibilidad de la estructura de las RNAs posibilitó la inclusión de variables dummies, relacionadas con la calidad de sitio y localización geográfica de los rodales (UBPFs), lo que permitió la obtención de óptimas capacidades de generalización, evidenciadas por los altos valores de R² y bajos valores de RMSE, obtenidas en las RNAs.
AGRADECIMIENTOS
Los autores del presente trabajo agradecen: a la Empresa Forestal Integral Macurije (Pinar del Río/Cuba), al Programa de Postgrado en Ciencias Forestales de la Universidad Federal Rural de Pernambuco (PPGCF/UFRPE), al Programa PEC-PG de CAPES Brasil y al Departamento Forestal de Universidad de Pinar del Río «Hermanos Saíz Montes de Oca».
REFERENCIAS BIBLIOGRÁFICAS
ALEIXO DA SILVA, J.A., 2015. Conceitos e princípios básicos de modelagem matemática em ciências florestais. Anais da Academia Pernambucana de Ciência Agronômica, vol. 11/12, pp. 195-215.
BIGLER, C. y BUGMANN, H., 2003. Growth-dependent tree mortality models based on tree rings. Canadian Journal of Forest Research, vol. 33 (2), pp. 210-221. DOI www.nrcresearchpress.com/doi/pdf/10.1139/x02-180.
BUCHMAN, R.G., PEDERSON, S.P. y WALTERS, N.R., 1983. A tree survival model with application to species of the Great Lakes region. Canadian Journal of Forest Research, vol. 13, pp. 601-608.
CAMPOS, J.C.C. y LEITE, H.G., 2017. Mensuração florestal: perguntas e respostas. 5a ed. S.l.: Viçosa: Editora UFV.
CHAPMAN, D.., 1961. Statistical problems in population dynamics. Proceedings of the fourth Berkeley symposium on mathematical statistics and probability. USA: Berkeley, CA, University of California Press, pp. 153-186.
CLUTTER, J.L. et al., 1983. Timber management: a quantitative approach [en línea]. 1sted. New York: John Wiley & Sons, 1983. Disponible en: https://www.amazon.com/Timber-Management-Quantitative-Jerome-Clutter/dp/0894647474
CLUTTER, J.L. y JONES, E.., 1980. Prediction of growth after thinning in old-field slash pine plantation [en línea]. S.l.: Res. Pap. SE-217. Asheville, NC: U.S. Dept. of Agriculture, Forest Service, Southeastern Forest Experiment Station. Disponible en: http://agris.fao.org/agris-search/search.do?recordID=US8044500
GLOVER, G.. y HOLL, J.., 1979. A basal area ratio predictor of loblolly pine plantation mortality. Forest Science, vol. 25 (2), pp. 275-282.
GUAN, BIING, T. y GERTNER, G., 1991. Modeling red pine tree survival with an artificial neural network. Forest Science, vol. 37 (5), pp. 14291440.
HAMILTON, D.A., 1974. Event probabilities estimated by regression [en línea]. INT-152. Utah, USA: USDA Forest Service, Intermountain Forest and Range Experiment Station, Ogden, Utah. Research Paper. Disponible en: https://www.biodiversitylibrary.org/bibliography/68708#/summary , https://doi.org/10.5962/bhl.title.68708 , https://www.biodiversitylibrary.org/bibliography/68708#/details.
HAMILTON, D.A., 1986. A logistic model of mortality in thinned and unthinned mixed conifer stands of Northern Idaho. Forest Science, vol. 32 (4), pp. 9891000.
HANN, D.W., 1980. Development and evaluation of an even- and uneven-aged ponderosa pine [en línea]. Arizona, USA: Arizona Fescue stand simulator. Ogden: Forest Service/Intermountain Forest and Range Experiment Station. Disponible en: https://archive.org/details/developmentevalu267hann.
K VANCLAY, J., 1994. Modelling forest growth and yield/ : applications to mixed tropical forests [en línea]. S.l.: Wallingford: CAB International UK. Disponible en: https://epubs.scu.edu.au/cgi/viewcontent.cgi?article=1538&context=esm_pubs.
L KING, S., BENNETT, K.P. y LIST, S., 2000. Modelling non-catastrophic individual tree mortality using logistic regression, neural networks, and support vector methods. Computers and Electronics in Agriculture, vol. 27, pp. 401-406.
LEE, Y., 1971. Predicting mortality for even-aged stands of lodgepole pine. Forestry Chronicle, vol. 47(1), pp. 29-32.
LORIMER, C.G., DAHIR, S.. y NORDHEIM, E.., 2001. Tree mortality and longevity in mature and old-growth hemlockhardwood forests. Journal of Ecology, vol. 89, pp. 960-971. DOI http://www.nrcresearchpress.com/doi/abs/10.1139/X10-14.
MALEKI, K. y KIVISTE, A., 2016. Individual tree mortality of silver birch (Betula pendula Roth) in Estonia. iForest, vol. 9, pp. 643-651.
PIENAAR, L.V. y SHIVER, B.D., 1981. Survival functions for site prepared slash pine plantations in the flat woods of Georgia Northern Florida. Southern Jour. Forestry, vol. 5 (2), pp. 59-62.
RICHARDS, F.J., 1959. A flexible growth function for empirical use. Journal of Experimental Botany, vol. 10, pp. 290-300.
SANQUETTA, C.R., 1990. Predição da sobrevivência em reflorestamentos de Pinus elliottii
Engelm [en línea]. Tesis de Maestría para la obtención del título de Máster en Ingeniería Forestal.
Brasil, Curitiba: Universidade Federal do Paraná. Disponible en:
https://www.bdpa.cnptia.embrapa.br/consulta/busca?b=ad&id=305301&biblioteca=vazio&busca=autoria:%
22SANQUETTA,%20%20C.%20R.%22&qFacets=autoria:%22SANQUETTA,%20%20C.%20R.%22&sort=&paginacao=t&paginaAtual=1.
SILVA, J.A.A. DA, 1986. Dynamics of stand structure in fertilized slash pine plantations [en línea]. Georgia Athens, USA: University of Georgia. Disponible en: https://books.google.com.cu/books/about/Dynamics_of_Stand_Structure_in_Fertilize.html?id=6Y5TNwAACAAJ&redir_esc=y.
VANCLAY, J.K., 1991. Mortality functions for north Queensland rain forests. Journal of Tropical Forest Science, vol. 4 (1), pp. 15-36.