
Efecto del grado de antropización en la estructura, en tres sitios fragmentados bosque siempreverde piemontano
Revista Cubana de Ciencias Forestales. 2017, 5(2):172-180
Efecto del grado de antropización en la estructura, en tres sitios fragmentados bosque siempreverde piemontano
Effect of the degree of anthropization in the structure, at three sites fragmented evergreen piedmont forest
Hugo Gabriel Sánchez Villacis1, Yudel García Quintana2, Gretel Geada López1, Yasiel Arteaga Crespo2, Jorge Rios Obregon2, Jessy Guerero Rubio2
1Ingeniero Agropecuario. Estudiante de Maestría. Universidad de Pinar del Rìo "Hermanos Saíz Montes de Oca".
Ecuador. Correo electrónico: ugosanch3z@hotmail.com ; gabriel@upr.edu.cu
2Doctor en Ciencias. Universidad Estatal Amazónica. Ecuador. Correo electrónico: yudelgarcia@gmail.com ; yarteaga@uea.edu.ec ; jrios@uea.edu.ec ; Jguerrero@uea.edu.ec
Recibido: 27 de marzo de 2017.
Aprobado: 13 de julio de 2017.
RESUMEN
La Amazonía ecuatoriana es reconocida a nivel mundial por su extraordinaria megadiversidad y la multiplicidad de bienes y servicios del bosque. Sin embargo, las prácticas inadecuadas de uso extractivo de los productos no maderables del bosque, la tala de extensas áreas de bosques para el desarrollo de la actividad petrolera y el aprovechamiento insostenido de madera como sustento económico de las comunidades han provocado cambios a nivel estructural y funcional en los ecosistemas. Se realizó el estudio en tres sitios de un bosque siempreverde, degradado del oriente amazónico (Mera, Shell y Puyo) con el objetivo de comparar el efecto del grado de antropización en la estructura, en tres sitios fragmentados bosque siempreverde piemontano. Se realizó un inventario florístico con 60 parcelas de 25 x 25 m2 y se midieron las especies arbóreas ≥ a 2.5 cm d1.30 y las especies en fase de regeneración natural con h < 2 m. Se encontraron 35 familias, 65 géneros, 101 especies y 2 298 individuos, siendo Arecaceae, Fabaceae y Moraceae las familias botánicas de mayor representación. El grado de antropización resultó altamente modificado, donde Mera fue el de mejor estado de conservación. Se evidenció una baja diversidad florística con patrones de alteración en la estructura vertical y horizontal, distinguido fitosociológicamente por dos estratos en los sitios de Shell y Puyo y por tres en Mera, indicador de cambios estructurales.
Palabras clave: Bosque siempreverde piemontano; estructura; grado de antropización; fragmentados.
ABSTRACT
The Ecuadorian Amazon is recognized worldwide for its extraordinary megadiversity and multiplicity of forest goods and services. However, the inadequate practices of extractive use of non-timber forest products, the clearing of extensive areas of forests for the development of oil activity and the unsustainable use of timber as economic sustenance of communities have led to structural and functional changes In ecosystems. The study was carried out in three sites of a degraded evergreen forest of the eastern Amazon (Mera, Shell and Puyo) in order to evaluate the effect of the degree of intervention on the forest structure. A floristic inventory was carried out with 60 plots of 25 x 25 m2 and tree species ≥ 2.5 cm d1.30 and species in natural regeneration phase with h <2 m were measured. We found 35 families, 65 genera, 101 species and 2 298 individuals, with Arecaceae, Fabaceae and Moraceae being the most representative botanical families. The degree of anthropization was highly modified where Mera was the best state of conservation. It was evidenced a low floristic diversity with patterns of alteration in the vertical and horizontal structure, distinguished phytosociologically by two strata in the sites of Shell and Puyo and by three in Mera, indicator of structural changes.
Key words: evergreen mousepads piedmont forest; structure; degree of anthropization; fragmented.
INTRODUCCIÓN
En la actualidad, se reconocen los esfuerzos a nivel global para la conservación de los bosques, expresados a través de convenios marcos, acuerdos mundiales y regionales en cada país, en los cuales se hace énfasis en la utilización sostenible de los componentes de la diversidad biológica, los productos forestales no maderables (PFNM) y el establecimiento de medidas correctivas en zonas degradadas, donde la diversidad biológica se ha reducido, así como el fomento de la cooperación entre autoridades y el sector privado para la aplicación de métodos científicamente fundamentados, orientados a la utilización sostenible de los recursos biológicos a escala local (Puerto Piñero, Raymond, & Condit S, 2014).
La riqueza ambiental, megadiversidad y, sobre todo, la realidad socioeconómica de las comunidades indígenas de Amazonía ecuatoriana requieren de una atención integral en la que se generen propuestas concretas para impulsar la conservación de los recursos forestales; por tradición, los campesinos en Ecuador aprovechan los PFNM de los bosques húmedos tropicales, bosques andinos, bosques secos, páramos y plantaciones forestales, para subsistencia y ocasionalmente para la venta (Aguirre Mendoza, 2013).
En los últimos años, Ecuador ha realizado varios esfuerzos para la valoración, caracterización de sus recursos, sin embargo, el conocimiento generado todavía no se está utilizando para la generación de políticas y toma de decisiones sobre el manejo. (Grijalva, et al., 2012).
La estructura de la vegetación, la diversidad de especies y los procesos de los ecosistemas forestales han sido identificados como componentes esenciales para la persistencia a largo plazo de los sistemas naturales, así como la regeneración natural de las especies sigue el mismo comportamiento de supervivencia manifestado por Espinosa et al., (2012). El conocimiento de las mismas proporciona información sobre aquellas especies más susceptibles a las perturbaciones en una región determinada y ayuda a predecir patrones sucesionales.
Aunque la Amazonía ecuatoriana es reconocida a nivel mundial por su extraordinaria megadiversidad y la multiplicidad de bienes y servicios del bosque; las prácticas inadecuadas de uso extractivo de los productos no maderables del bosque, la tala de extensas áreas de bosques para el desarrollo de la actividad petrolera y el aprovechamiento insostenido de madera como sustento económico de las comunidades han provocado cambios a nivel estructural y funcional en los bosques siempreverdes.
El objetivo de este trabajo fue comparar el efecto del grado de antropización en la estructura, en tres sitios fragmentados bosque siempreverde piemontano..
MATERIALES Y METODOS
Ubicación del área de estudio
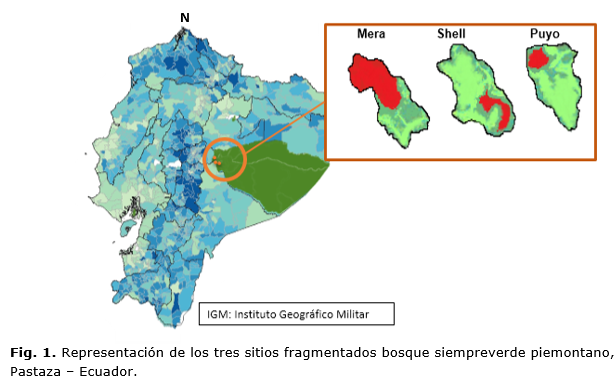
Grado de perturbación
Se evaluó el grado de perturbación mediante el método de observación, a partir de los criterios establecidos por Matos y Ballate (2006). Se estableció el criterio de bajo, moderado y alto, considerado bajo aquella área que resulte con poca o casi ninguna alteración (menos del 5 %), moderada cerca de un 50 % de afectaciones producidas por el hombre y alto cuando se aprecie alta influencia antrópica (más del 80 %).
Se realizó un análisis de correspondencia para determinar la relación entre las variables nominales (grado de antropización) y los sitios. Se determinó la dimensionalidad de la solución a partir del mínimo ([filas, columnas] -1). Para ello, se empleó el paquete estadístico SPSS ver. 21.0.
Inventario florístico
Se realizó un inventario de la vegetación leñosa con diámetros mayores e iguales a 2,5 cm. (d1.30 ≥ 2,5 cm.) y alturas mayores e iguales a 2,0 m. (h < 2 m.) mediante el recuento físico por especie en 60 parcelas distribuidas, 20 en cada sitio, las cuales fueron georreferenciadas con la utilización de un GPS RTK precisión de 1 cm. El inventario se realizó por el método de área mínima con parcelas cuadradas de 25 x 25 m2 mediante muestreo sistemático. Se hizo la identificación botánica preliminar en el campo y se confirmó con Catálogo de Plantas Vasculares del Ecuador (Jorgensen y León, 1999), Libro Rojo de las Plantas Endémicas del Ecuador (1) y constatada con la colección de muestras del Herbario Ecuatoriano Amazónico (ECUAMZ).
Estructura y composición florística
Se calculó el índice de valor de importancia ecológica (IVI) mediante lo descrito por Lamprecht (1990) para todas las especies con diámetros superiores a 2,5 cm.
Se determinó la diversidad florística mediante los índices de Shannon, Simpson y Equidad de Hill (Moreno, 2001) con la ayuda del software Biodiversity Professional. Se realizó un análisis de varianza de clasificación simple con los valores de diversidad entre los sitios con un nivel de confiabilidad del 95 %. Se clasificaron las unidades de muestreo de acuerdo con la composición florística mediante análisis de conglomerados jerárquicos, a partir del índice de similitud de Sorensen.
RESULTADOS Y DISCUSIÓN
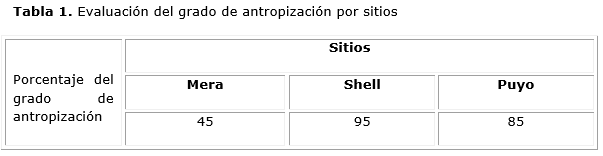
En la tabla 1, se muestra la evaluación del grado de antropización de los sitios de estudio donde Mera resultó en la categoría moderado (45 %), mientras que la Shell y Puyo resultaron con alta antropización (más del 80 %). Estos resultados obedecen a las actividades antropogénicas que se realizan de manera sistemática en los sitios de estudio.
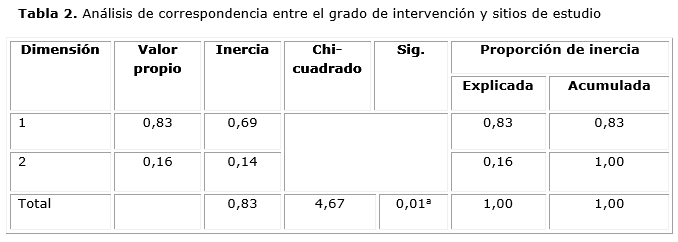
El análisis de correspondencia mostró una correlación significativa (p ≤ 0,05) entre los sitios de estudio y el grado de antropización (Tabla 2) con valores de inercia total de 0,83. La solución indicó que solo es significativo el primer eje con una proporción de inercia que explicó el 83,5 % de la varianza total. Este resultado demostró el alto grado de deterioro de las áreas de bosque donde se realizan las actividades de tala y desmonte, considerados los más vulnerables ante las perturbaciones.
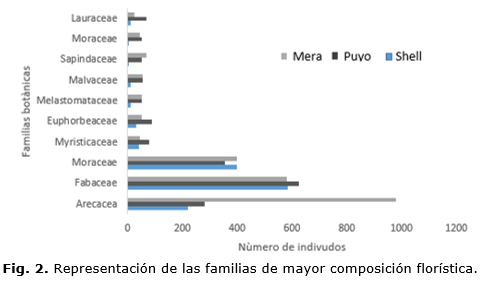
En la figura 1, se aprecian las diez familias botánicas de mayor representación en cada sitio de estudio. El inventario florístico en el sitio Mera resultó superior e incluyó un total de 35 familias, 65 géneros, 101 especies y 1259 individuos, resultando con mayor riqueza en orden representativo: Arecaceae, Fabaceae y Moraceae. La riqueza a nivel familiar encontrada en este sitio coincide con lo reportado por Gentry (1982), Yaguanoet al., (2012), donde refieren que estas familias constituyen elementos distintivos del bosque húmedo tropical, caracterizados por una flora entre la vegetación de tierras bajas y de cordillera.
La flora descrita para la Shell incluyó un total de 12 familias botánicas, 24 géneros, 30 especies y 958 individuos, mientras que para el sitio Puyo los resultados fueron similares aunque con valores discretamente superiores en términos de riqueza, con un total de 16 familias botánicas, 26 géneros, 32 especies y 746 individuos, lo cual pudiera deberse a las prácticas de tala sin criterios silvícolas con fines de explotación petrolera y uso maderable y no maderable como sustento económico de los colonos, lo que ocasiona cambios en la composición florística de la flora amazónica.
Un elemento importante a resaltar es la poca representación de individuos por cada especie. Estos resultados coinciden con lo reportado por Lamprecht (1990), los cuales refieren que una de las características particulares de los bosques tropicales es el gran número de especies representadas por pocos individuos con patrones complejos de tipo espacial entre el suelo y el dosel. En los sitios Shell y Puyo, esta característica está menos atribuida que en Mera, lo que sugiere un bosque con tendencia a la homogeneidad por el predomino de pocas especies, debido principalmente a la intervención antrópica por las prácticas de extracción.
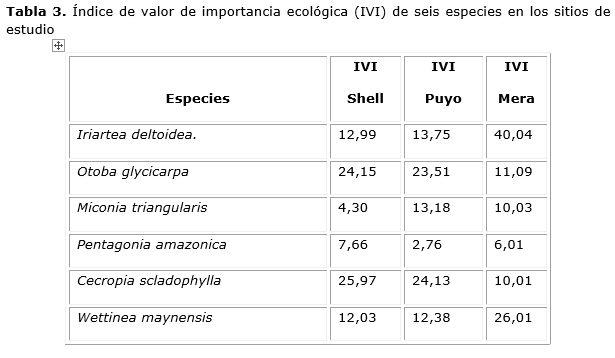
El índice de valor de importancia ecológica de las seis especies que resultaron con d1,30 > 2,5 cm. del sitio de Mera resultó superior en relación con los sitios Shell y Puyo y en la mayoría de los casos las especies más abundantes se tornaron menos frecuentes y viceversa (Tabla 3). Entre las especies de mayor peso ecológico, Iriarteadeltoidea Ruiz & Pav, Otoba glycycarpa (Ducke ,Rodrigues& T.S. Jaram), Cecropiasciadophylla (Mart) y Wettineamaynensis(O.F.Cook& Doyle)(36,51 % de la flora) ocuparon las primeras posiciones ecológicas, fundamentalmente por su abundancia relativa, acumulando de conjunto el 50 % del total de valor de importancia, aunque es notorio resaltar que Irialtadeltoide ocupó una posición alta en el sitio de Mera por su dominancia, dada el área basal, pero resultó poco abundante.
Las especies menos representadas, según Moreno (2001), pueden ser más sensibles a las perturbaciones ambientales y alerta acerca de procesos empobrecedores. Estos resultados revelan que las especies que dominan la comunidad, en estos momentos, son típicas de bosques secundarios degradados, debido al impacto de las talas fundamentalmente. Este proceso ha favorecido la proliferación de otras especies con capacidad de tolerar las condiciones ecológicas de los bosques siempreverdes amazónicos.
Es notorio resaltar los cambios que se dan en la importancia ecológica de las primeras seis especies en cada sitio, lo cual es un indicador de cambios estructurales producto de las consecuentes talas que se realizan, sobre todo, en los sitios de la Shell y Puyo.
En Mera, la distribución por clases diamétricas indicó que el 71,13 % del total de individuos se concentró en las primeras clases (2,5 - 22,5 cm.), el 20,05 %, en la segunda clase (22,6 - 42,6 cm.), mientras que solo un 8,82 % de los individuos ocuparon las clases superiores (42,5 - 62,5 cm.) (Figura 2). En los sitios Shell y Puyo, la distribución por clases diamétricas resultó inferior a Mera en todas las clases diamétricas, con una estructura más heterogénea. Se obtuvo que el 39,79 % del total de individuos se concentró en las primeras clases (2,5 - 22,5 cm.), el 66,25 %, en la segunda clase (22,6 – 42,6 cm), mientras que solo un 3,96 % de los individuos ocuparon las clases diamétricas superiores (42,5 - 62,5 cm.).
Según Lamprecht (1990), la mayoría de las áreas naturales existentes presentan una estructura disetánea, caracterizada por diferentes edades y ritmos de crecimiento; de ahí que las diferencias existentes entre sitios obligan a establecer una tipificación para el manejo adecuado de los bosques. De acuerdo con estos resultados, se puede afirmar que todos los sitios han sido perturbados por la tala intensiva de los individuos de mayor diámetro, con mayor énfasis en Shell y Puyo donde se extraen altos volúmenes de madera, lo cual ha alterado la estructura diamétrica de este bosque con predominio de árboles con diámetros más pequeños y una estructura irregular.
La forma de una distribución gráfica similar a una “J” invertida, predominante en el sitio Mera, es característico de los bosques tropicales heterogéneos y disetáneos, con tendencia a disminuir en número de individuos a medida que va aumentando el diámetro. Estos resultados coinciden con (Escobar, 2013) en un inventario realizado en áreas asociadas a actividades agropecuarias en el cerro Quinini, Colombia.
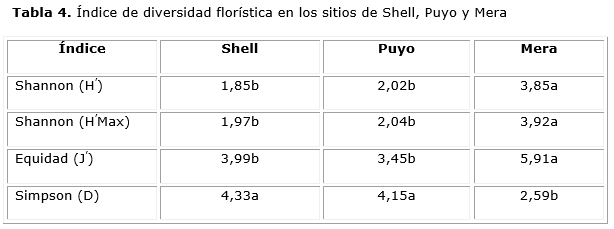
La diversidad florística resultó baja para los sitios de estudio (Tabla 4), significativamente inferior en Shell y Puyo (p ≤ 0,05). El sitio de Mera se presentó como el área más diversa, expresado a través del índice de Shannon, con mayor equitatividad (Equidad) y menor dominancia (Simpson). Estos resultados se corresponden con lo reportado por Samaniego et al., (2015) atribuyendo la baja diversidad a la influencia de la antropización.
Según Albis (2009), los bosques naturales tropicales son heterogéneos y están conformados por una alta diversidad de especies, con diferentes edades, diferenciados en tres estados sucesionales: brinzal, latizal y fustal. En las áreas de estudio, esto no se corresponde por las perturbaciones que limitan e interrumpen los estados sucesionales del bosque.
CONCLUSIONES
Se comprobó que el estado actual del bosque siempreverde piemontano de los sitios Mera es ciertamente desfavorable con un porcentaje menor en comparación a Puyo y Shell, obteniendo un efecto significativamente más marcado en estos últimos, determinado por las prácticas de tala sin criterios silvícolas, lo que influyó en la baja diversidad florística.
Se constató cambios y alteraciones en la estructura vertical y horizontal del bosque con predominio de las familias botánicas Arecaceae, Fabaceae y Moraceae y se notó escaso valor de importancia ecológica en especies características de esta formación boscosa.
La estructura del bosque resultó heterogénea con alteraciones en las clases diamétricas, predominio de individuos en las clases inferiores y una distribución en forma de J invertida para Mera y de campana de Gauss para Shell y Puyo como indicador del estado de sucesión ecológica.
REFERENCIAS BIBLIOGRÁFICAS
León-Yánez, S. R. Valencia, N. Pitman, L. Endara, C. Ulloa Ulloa& H. Navarrete. Libro Rojo de las Plantas Endémicas del Ecuador, Segunda edición. Publicaciones del Herbario QCA. Quito. : Universidad Católica del Ecuador, 595 P, 2012.
Ordóñez, Thelma maría mejía. Composición y estructura florística en fragmentos de bosques en los municipios de Yuscarán, Oropolí y Güinope, Departamento de el Paraíso, Honduras, c.a.Honduras: Revista ciencias espaciales, volumen 6, número 1 primavera, 2013.
Grijalva, J., X. Checa, R. Ramos, P. Barrera y R. Limongi. Situación de los Recursos Genéticos Forestales – Informe País Ecuador. Preparado por el Programa Nacional de Forestería del INIAP con aval del INIAP/FAO/MAE/MAGAP/MMRREE. Documento sometido. 2012.
Escobar, N. Diagnóstico de la composición florística asociada a actividades agropecuarias en el cerro Quinini. Colombia: Universidad de Cundinamarca (sede Fusagasugá) facultad de ciencias agropecuarias - grupo de investigación área verde, 2013.
Puerto Piñero, Carolina, Raymond, E y Condit S, Richarrd. Metodologías para el sistema de monitoreo de la diversidad biológica de Panamá. Panamá: s.n., 2014.
Espinosa, C.I., De la Cruz, M., Luzuriaga, A. L. y Escudero. Bosques tropicales secos de la región Pacífico Ecuatorial: diversidad, estructura, funcionamiento e implicaciones para la conservación. S.l.: Ecosistemas 21 (1-2): 167-179, 2012.ISBN 1697-2473.
Matos, J. y Ballate, D. ABC de la Restauración Ecológica. Santa Clara: Editorial Feijóo. 92 p., 2006.
Moreno, C. Métodos para medir la biodiversidad. Manuales y Tesis. Zaragoza, España. 84 p: SEA. Volumen 1, 2001.
Lamprecht, H. Los ecosistemas forestales en los bosques tropicales y sus especies arbóreas-posibilidades y métodos para un aprovechamiento sostenido. Alemania. 335 p. Cooperación técnica. Eschborn, 1990.
Composición florística y estructura de los bosques secos y su gestión para el desarrollo de la provincia de Loja, Ecuador. Aguirre Mendoza, Zhofre, y otros. Cuba: Revista Científica Avances, 2013, Vol. Vol. 15 .ISSN 1562-3297.