
Caracterización del diámetro, ángulo de inserción y longevidad de ramas vivas axiales de Nothofagus obliqua
Revista Cubana de Ciencias Forestales. 2017, 5(2):127-139
Caracterización del diámetro, ángulo de inserción y longevidad de ramas vivas axiales de Nothofagus obliqua
Characterization of the diameter, branch angle and longevity of axial branches of Nothofagus obliqua
Patricio Corvalán Vera
Ingeniero Forestal. Máster en Ciencias Forestales. Universidad de Chile. Facultad Ciencia Forestales y de la Conservación de la Naturaleza. Departamento Gestión Forestal y de su Medio Ambiente, Universidad de Chile. Correo electrónico: pcorvalanvera@gmail.com
Recibido: 27 de marzo de 2017.
Aprobado: 13 de julio de 2017.
RESUMEN
El desconocimiento de la dinámica de crecimiento de la copa viva en los renovales de la especie Nothofagus obliqua limita fuertemente la objetiva formulación de esquemas silvícolas orientados a la producción de madera industrial de alta calidad. Por ello en este trabajo se describieron relaciones entre el tamaño, ángulo de inserción y edad de las ramas en el perfil fustal y algunas variables de la copa y del árbol. A una muestra de 59 árboles dominantes, distribuidos en distintas condiciones de edad, se le aplicó una técnica combinada de análisis de tallo, función de ahusamiento y mediciones de la rama más gruesa en cada decil de la altura total. Esto permitió determinar significativas correlaciones entre el diámetro, ángulo de inserción y la edad de la rama, así como con el tamaño y la edad de los árboles. Asimismo, las ramas más gruesas tendieron a tener ángulos de inserción menores, mayor edad, estar ubicadas en alturas relativas más bajas y estar insertas en secciones fustales de mayor diámetro. A partir de estas relaciones se posibilita la formulación de futuros modelos de predicción de ramas como herramientas para el desarrollo de esquemas silvícolas que se adapten a los diferentes requerimientos de calidad de la madera.
Palabras clave: arquitectura del árbol; luminosidad; tamaño; ángulo de inserción y edad de ramas.
ABSTRACT
The lack of knowledge about grow dynamics of the living tree crown of Nothofagus obliqua secondary growth forests strongly limits the objective formulation of silvicultural schemes oriented to the industrial production of high quality wood. Therefore, in this work, we described basic relationships between tree size, age and angle branches insertion and the crown. Considering a sample data of 59 dominant trees, distributed in different age conditions, we applied a combined analysis technique of stem analysis, steam taper analysis and thickest branch measurement in each decile of the total height. This approach allowed us to determine that there is a significant relationship between the steam diameter, the angle insertion and the age of the branch, as well as the size and age of the trees. Also, the thicker branches tend to have lower insertion angles, to be older, to be located at lower relative heights and to be located in larger diameter sections. Taking into consideration these relationships, it is possible to build new predicted branch models as tools for the development of silvicultural schemes to suit different log grade.
Key words: branch; size; angle and age of branches; luminosity; tree architectural.
INTRODUCCIÓN
Las ramas son estructuras del árbol que cumplen funciones de soporte y conductividad hidráulica entre las raíces, el fuste y el follaje activo fotosintéticamente (Garber y Maguire, 2005). La vitalidad de la rama está fuertemente controlada por la luminosidad a la que está expuesto el follaje que la soporta (Maguire et al. 1991, Roeh y Maguire 1997). La dinámica de formación del tejido leñoso permite al árbol una permanente agregación, adecuación y expansión de las ramas en búsqueda de luminosidad, dejando tras de sí una estructura vertical de copa viva y muerta como resultado de su autoconstrucción y competencia con el entorno (Ford 1985, Kozlowski y Pallardy 1997, Lowell et al. 2014). La configuración espacial, así como el tamaño y la forma de las ramas vivas y muertas, tiene mucha importancia en la formación de los nudos en la madera (Maguire et al. 1991, Roeh y Maguire 1997, Trincado y Burkhart 2008). Los nudos limitan la calidad de la madera que se produce en el segmento del fuste al cual están adheridos. Es especialmente importante el diámetro y el ángulo de inserción de la rama para la determinación del aprovechamiento y el valor comercial de la madera para debobinado y aserrío, por razones de deterioro estético, de resistencia y de desvío del grano que presentan los nudos (Larson et al. 2001, Trincado y Burkhart 2008, Lowell et al. 2014).
El control de la luminosidad de la copa mediante los tratamientos silviculturales es esencial en la producción de madera de alta calidad ya que estos deben realizarse en la etapa temprana de formación y agregación de tejido basal del fuste. Raleos y podas tardíos tendrán escaso o nulo efecto económico sobre la posición basal del fuste ya que solo afectará a la madera perimetral (Kozlowski y Pallardy 1997, Larson et al. 2001). Para la comprensión de la dinámica en la formación de nudos, es fundamental conocer el proceso natural de desarrollo de las ramas vivas a lo largo del perfil fustal a fines de dimensionar la envergadura del problema de la presencia de nudos (Roeh y Maguire 1997).
Diversos estudios relacionan características morfológicas y arquitecturales de ramas del género Nothofagus con la luminosidad, gradiente altitudinal y ontogénesis, destacándose los trabajos desarrollados en Nothofagus alpina (Poepp. et Endl.) Oerst., Nothofagus obliqua (Mirb.) Oerst. y Nothofagus pumilio (Poepp. et Endl.) Krasser. (Read y Hill 1985, Puntieri et al. 2013). Para N. obliqua, las principales características que se reportan son que en bosques sombreados las ramas axiales son más delgadas, cortas y con menos hojas y ramas secundarias que en bosques más asoleados. También se reporta que la especie tiende a ramificarse en forma menos jerárquica y a perder vigor bajo sombra.
Los renovales de N. obliqua en Chile, originados por la conversión de los bosques primarios (Donoso, 1993) han sido escasamente intervenidos silvícolamente con fines de producción de madera de alta calidad y tienen un alto potencial productivo (Grosse y Quiroz 1999). Los incentivos para el manejo, originados en la dictación de la ley 20.283 de Fomento a la Protección y Manejo del bosque nativo, abren un interesante espacio para iniciar la silvicultura de renovales con fines de producción de madera de alta calidad. Los principales trabajos realizados en silvicultura de renovales de roble en Chile han estado orientados a definir las tasas de crecimiento como respuesta ante distintas intensidades de raleo, sin embargo, existe un gran desconocimiento sobre el tamaño de las ramas en el perfil fustal. Son escasos los trabajos que dan cuenta del efecto de la luminosidad en el tamaño de las ramas (Gspaltl et al. 2013, Corvalán 2015).
La hipótesis de trabajo asume que la vitalidad de las ramas de N. obliqua está controlada por la luminosidad que recibe durante su formación; esto es, 1) las ramas más bajas sometidas a fuerte competencia inicial tendrán mayor probabilidad de presentar ángulos de inserción pequeños que a alturas superiores, 2) que estarán mayor tiempo activas y, por lo tanto, 3) tendrán mayor desarrollo diamétrico que las ramas superiores, 4) que se encontrarán insertas en secciones fustales de mayor edad y que, análogamente, 5) estas características se verán acentuadas por la edad y tamaño de los árboles. Considerando la importancia económica que los árboles dominantes tienen en el retorno final de las operaciones forestales (Ford, 1985), en este trabajo, se cuantifica y describe la relación existente entre el tamaño, ángulo de inserción y edad de las ramas vivas de mayor diámetro en el perfil de los árboles pertenecientes al cuartil superior del diámetro normal a 1,3 o diámetro a la altura de pecho (DAP) en rodales no manejados, sujetos a autoraleo, no descritas a la fecha..
MATERIALES Y METODOS
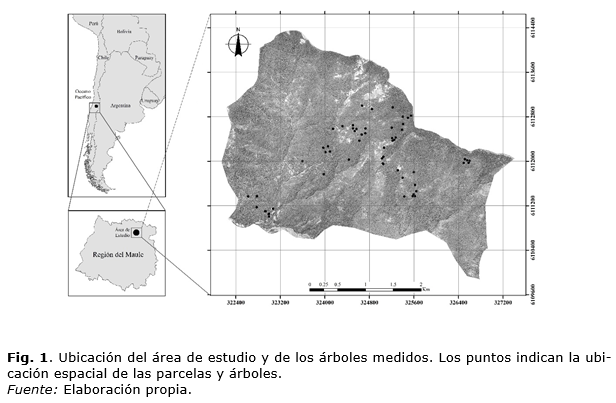
Área de estudio
El área de estudio se ubicó a 31 km., al este de la ciudad de Molina en los contrafuertes cordilleranos donde termina el valle del estero Upeo (Chile) (35° 07’ S, 70° 56’ O; 698 msnm.) cubriendo aproximadamente 1.300 hectáreas. El área presenta predominantemente una exposición sur y una topografía abrupta con mesetas interiores (Figura 1). Los suelos corresponden a la asociación Sierra Bellavista, Mollic Haploxeralf (Alfisol), desarrollados a partir de materiales volcánicos (andesita) y en algunos sectores recubiertos por cenizas volcánicas frescas en pendientes de 30% a 50%. El clima mediterráneo presenta en el área una temperatura media anual de 12,3 ºC, una precipitación anual de 1.315 mm año-1, con un período libre de heladas de 216 días al año y una humedad relativa media del 68%. La vegetación natural corresponde a la región del bosque caducifolio de La Montaña, que por el contrafuerte cordillerano andino se distribuye desde el sur de la región del Libertador Bernardo O'Higgins hasta el norte del río Biobío (Gajardo, 1994). Esta formación vegetal se caracteriza por su gran riqueza florística y ser el límite norte de algunas especies leñosas y herbáceas más características de los bosques australes. Las comunidades características de esta formación son Nothofagus obliqua-Ribes punctatum Ruiz et Pav. y Austrocedrus chilensis (D. Don) Pic. Serm. et Bizzarri (Gajardo, 1994). La zona de estudio se inserta en el piso de vegetación denominado "Bosque caducifolio mediterráneo andino de Nothofagus obliqua y Austrocedrus chilensis". Este piso de vegetación se distribuye en laderas medias de la Cordillera de Los Andes de la región del Maule y norte del Biobío, entre los 900 y 1.800 msnm. Corresponde a un bosque caducifolio con participación de ciprés de la cordillera Austrocedrus chilensis y especies esclerófilas, principalmente Lithrea caustica (Molina) Hook. et Arn. y Quillaja saponaria Mol (Luebert y Pliscoff 2006).
Selección de árboles y medición de ramas
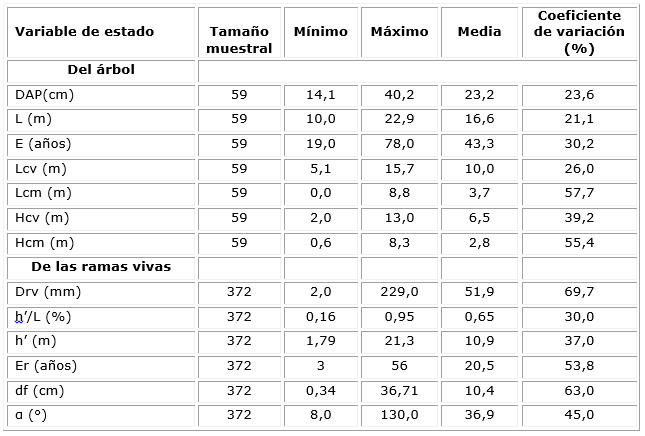
Los rodales de N. obliqua sin manejo, en los cuales se seleccionaron los árboles para el presente estudio fueron elegidos cubriendo distintos estados de desarrollo requeridos para la construcción de un diagrama de manejo de la densidad (Corvalán, 2015). Las variables dendrométricas se midieron en 59 parcelas: 41 circulares de 200 m2 y 18 de tamaño y forma variable. Las condiciones ambientales de la muestra corresponden a las que la ley permite para su intervención (menores a 60% de pendiente y estar fuera de los cursos de agua). Los criterios de selección de tamaños y formas de las parcelas se describen detalladamente en Corvalán (2015). La ubicación espacial de las parcelas se presenta en la figura 1. De cada parcela, se seleccionó el árbol central del cuartil superior de la distribución diamétrica lo que representa la fracción del rodal de mayor interés comercial. Para cada uno de los 59 árboles, se midió el (DAP, cm). Luego se volteó y se realizaron las siguientes mediciones: largo total (L, m), largo de copa viva (Lcv, m) y largo de copa muerta (Lcm, m), a partir de las cuales, por sustracción del largo total, se derivaron la altura de inicio de copa viva (Hcv, m) y la altura de inicio de la copa muerta (Hcm, m). El fuste fue dividido en 10 deciles de altura donde se identificó y midió en cada uno la rama axial de mayor diámetro en dirección perpendicular a su eje. El diámetro fue medido a 1 cm. de distancia desde el punto de inserción en el fuste (h, m) para evitar la protuberancia axial de la rama. Se clasificó cada rama en viva o muerta y se midió su diámetro de ramas vivas y muertas (Drv, mm; Drm, mm) y su ángulo de inserción superior en el fuste (α, °). Desde la base, se midió el diámetro fustal con corteza (df, cm) cada un metro aproximadamente para evitar las irregularidades fustales. Se realizó un análisis de tallo y se extrajeron discos fustales cada un metro aproximadamente, con excepción de pocos árboles donde fue de mayor longitud (2,40 m).
Estimación de la edad del árbol y de las ramas
Para calcular la edad de las ramas (Er, años), se consideró que su año de formación, a la altura de inserción en el fuste (h’, m) corresponde al año cronológico en que la médula del fuste y el de la rama se unen. Por ello, se utilizó análisis de tallo y la interpolación de alturas interiores totales. La altura de inserción de la rama en el fuste (h´) se estimó a través de una relación trigonométrica que corrige el punto de emergencia de la rama en altura (h), de acuerdo con su ángulo de inserción (α) como se indica en la ecuación [1]. La edad del árbol (E, años) se estimó contando los anillos anuales del disco fustal a la altura del tocón. El radio del fuste (df, cm.) en el punto de emergencia de la rama se calculó usando una regresión polinómica de sexto grado, a partir de los datos de ahusamiento del árbol. El método de estimación de la edad de la rama se presenta en Corvalán (2015).
 .
.
RESULTADOS
Dimensión de las copas
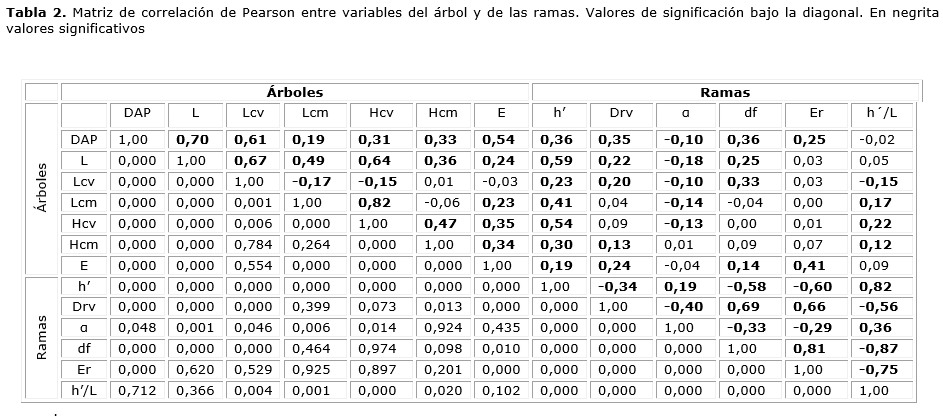
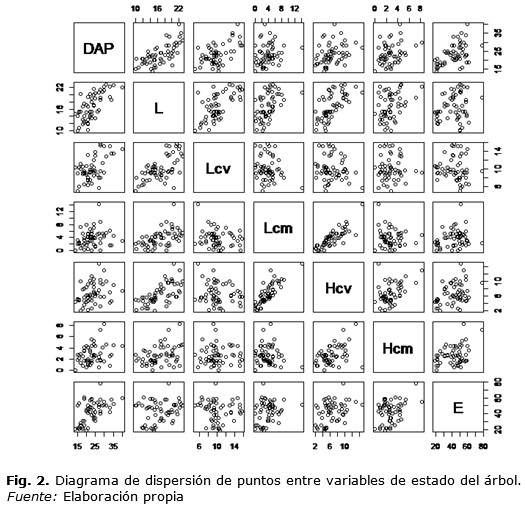
Tal como se señala en la Tabla 1, el largo de la copa viva en promedio cubrió el 60,2 % del largo total con un inicio de copa a los 6,5 metros, extendiéndose por 10 m. en promedio y con una variación de 26 %. Lcv se relacionó significativamente con el tamaño del árbol (DAP y L) con correlaciones de 0,61 y 0,67 respectivamente (Tabla 2 y figura 2). La copa muerta cubrió el 22,3 % del total, se inició a los 2,8 m., se extendió por 3,7 m. y presentó una alta variabilidad. La extensión de la copa muerta se relacionó significativamente con la altura de inicio de la copa viva y, en menor medida, con el largo total del fuste. Lcv se relacionó significativamente con el Lcm con correlación de -0,17. Lcv, en promedio, se inició a los 6,5 m., con una variación del 39,2 % y se relacionó muy estrecha y positivamente con el largo total del fuste. A su vez, Lcm se inició en promedio a los 2,8 m., con una variación del 55,4 %. La altura de inicio de la copa muerta se relacionó significativamente con la altura de inicio de la copa viva y el tamaño del árbol (DAP, L). Tanto la altura de inicio de copa viva como el de copa muerta se relacionaron significativamente con la edad del árbol con coeficientes de correlación de 0,35 y 0,34.
Tabla 1. Estadísticas descriptivas de la muestra.
Diámetro de las ramas
Considerando el total de ramas vivas en todo el perfil del árbol, el diámetro promedio fue de 51,9 mm., con valores extremos de 2,0 mm. y 229,0 mm. Como se muestra en la figura 3, el tamaño se explica por la posición relativa de las ramas en altura total. El Drv presentó grandes variaciones en el perfil de alturas relativas, donde tuvo su máxima amplitud en la porción basal, convergiendo hacia un valor intermedio hacia el 60% en h´/L y, desde allí hasta el ápice, decreciendo hasta llegar a cero. La gran dispersión de diámetros de la rama viva también se explicó por el diámetro fustal en su punto de inserción y la edad de la rama con las que se correlacionó con valores significativos de 0,69 y 0,66, respectivamente (Tabla 2 y figura 3). Otro factor determinante en Drv fue α, con la que se relaciona significativamente con correlación de -0,40. Drv se correlacionó positivamente con el DAP y la edad del árbol con valores de 0,35 y 0,24.
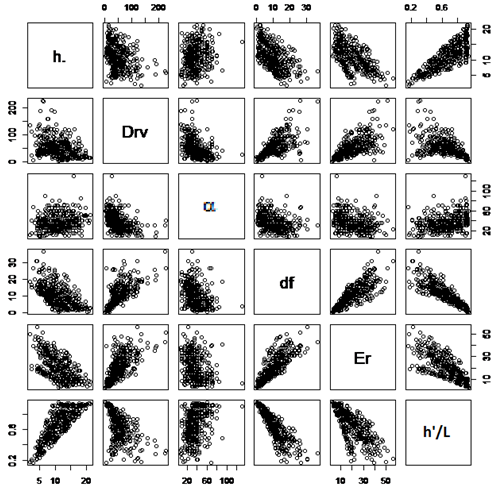
Fig. 3. Diagrama de dispersión de puntos entre variables de estado de las ramas.
Fuente: Elaboración propia.
Ángulo de las ramas
El ángulo de inserción de las ramas vivas presentó un monto promedio de 36,9° con valores extremos de 8° y 130° y se relacionó significativamente con el diámetro del fuste y la edad de la rama con correlaciones de -0,33 y -0,29. Contrariamente, el ángulo aumentó con la altura relativa con la que alcanzó una correlación de 0,36. El ángulo α se relacionó negativamente con el tamaño del árbol y el largo de copa viva y muerta; no se relacionó con la edad del árbol.
Edad de las ramas
La edad de las ramas vivas en la muestra tuvo en promedio 20,5 años con extremos de 3 y 56 años y un coeficiente de variación de 53,8 %. Estos valores estuvieron significativamente ligados al diámetro del fuste en su punto de inserción y al diámetro de la rama con correlación de 0,81 y 0,66 respectivamente; se relacionaron significativamente con la altura relativa de inserción con correlación de -0,75. La edad de las ramas se asoció significativamente con la edad del árbol y el DAP con correlaciones de 0,41 y 0,25, respectivamente.
DISCUSIÓN
La muestra dendrométrica temporal utilizada en este trabajo permite establecer algunas relaciones básicas entre el tamaño, ángulo de inserción y edad de las ramas vivas y algunas variables de tamaño, edad y dimensiones de la copa del árbol que presentan los árboles dominantes en renovales de N. obliqua, no sometidos a manejo. La muestra cubre diversos estados de desarrollo y corresponden a los estados se inicia el manejo forestal (Corvalán, 2015). La figura 2 señala con claridad cómo la copa viva y muerta se extiende proporcionalmente al desarrollo en altura total del árbol, coincidiendo con Changsheng et al. (1998) quienes muestran proporciones similares de copa viva en Picea abies (L.) Karst., una especie europea. La longitud de la copa viva está fuertemente ligada a la competencia, la que es más pequeña en condiciones de alta densidad (Mäkinen 2002, Kantola y Mäkelä 2004). Kantola y Mäkelä (2004), estudiando el desarrollo de la copa en P. abies, concluyen que la razón de copa decrece con la edad del árbol y la densidad, al comparar rodales raleados con no raleados. El efecto de la competencia sobre la copa viva parece notable al comparar, en latifoliadas, árboles creciendo aislados que cubren entre el 39 % y 60 % (Russell y Weiskittel, 2011), con árboles en bosques densos y raleados donde cubren entre el 27% y 38% (Sonderman, 1984). La positiva relación que se encontró de la extensión de la copa muerta con la altura de inicio de la copa viva y el largo fustal resulta análoga a lo reportado por varios autores que dan cuenta de la gradualidad del receso de la copa.
El tamaño promedio que alcanzaron las ramas vivas de mayor tamaño de N. obliqua fue muy superior al rango que Groot y Schneider (2011) establecen para especies de hoja ancha en Canadá, superiores a los valores máximos reportados por Garber y Maguire (2005) en coníferas en Oregon, aun creciendo en bajas densidades. En relación con la distribución del diámetro de las ramas en el perfil vertical de alturas que se muestra en la figura 3, es concordante con la señalada en otros estudios, donde se observan diámetros pequeños en la base y la punta del árbol, existiendo una posición intermedia donde es posible encontrar valores máximos que dependen de la especie, densidad y la edad (Colin y Houllier 1992, Garber y Maguire, 2005). La positiva relación que se encontró entre el diámetro y la edad de la rama no coincide plenamente con lo señalado por Mäkinen (2002) quien, estudiando el efecto de la densidad de Betula pendula Roth en Finlandia encuentra que el incremento en diámetro de las ramas tiene un máximo, lo cual decrece posteriormente. Esta discrepancia se puede deber a que Mäkinen (2002) estudia las ramas vivas y muertas en todo el perfil fustal y en este trabajo solo se consideraron las ramas vivas. Garber y Maguire (2005), estudiando árboles en bosques mixtos en Oregon, señalan que el tamaño de las ramas depende de su posición en la copa, la posición social del árbol y la estructura del rodal. En este trabajo, al igual que Mäkinen y Colin (1999) encuentran que el diámetro de la rama está positivamente correlacionado con el diámetro del fuste. Al comparar el diámetro de ramas axiales de este estudio, con los reportados por Puntieri et al. (2013) en árboles juveniles-adultos de N. obliqua en el sur de Argentina, se advierte una notable diferencia de tamaños, siendo estos últimos del orden de 6 a 19 cm., en comparación a los 51,9 cm., del presente estudio.
Esta diferencia no se explica por la posición de las ramas, ya que aquellas provienen de los primeros metros y de las ramas de mayor diámetro del árbol; ni tampoco, por la luminosidad, ya que ambos estudios cubren todo el rango. Las discrepancias se pueden explicar por la diferencia de edad de la muestra (23 a 29 años contra 43 años en este estudio) o bien por las diferencias de latitud entre ambos estudios, representando los extremos de la distribución de esta especie.
Existen escasos estudios del ángulo de inserción de ramas en latifoliadas (Nelson et al. 2014). El ángulo α encontrado en este estudio resulta tener un valor promedio similar a varios álamos y abedules en Maine (Nelson et al. 2014); pero muy inferior al reportado por Roeh y Maguire (1997) para plantaciones juveniles de Pseudotsuga menziesii (Mirb.) Franco, es decir, ramas más agudas. La tendencia de disminución del ángulo de inserción de la rama con la profundidad de copa encontrada difiere notoriamente con la que reportan en plantaciones Roeh y Maguire (1997) en P. menziesii. Esta discrepancia puede deberse a las diferentes tolerancias a la luz que presenta el follaje de N. obliqua y P. menziesii. La mayor demanda de luz y el alto nivel de competencia inicial por la dominancia apical que se producen en los densos vástagos de la regeneración de roble pueden estar explicando parte de las diferencias. De acuerdo con Larson et al. (2001), el ángulo de inserción de las ramas en Pinus radiata D. Don está negativamente relacionado con el diámetro de la rama en concordancia con el presente estudio. También señalan que esta variable está bajo fuerte control genético. Nelson et al. (2014) destacan que la forma de las copas en maderas duras está a menudo compuesta con ángulos verticales menores que las de coníferas. El ángulo promedio de ramas axiales encontrado en este estudio de 36,9° resulta muy inferior al encontrado por Puntieri et al. (2013) 62° para la misma especie, sin embargo, el rango resulta muy similar (8° a 130°, en este estudio, contra 0° a 140°). Las razones que explican la diferencia entre los promedios puede ser la misma que para lo diámetros: diferencias de edad y latitud.
El valor máximo de 56 años de las ramas vivas, reportado en este estudio, resulta mucho mayor que las ramas vivas basales de la copa viva de 21, 17 y 15 años encontradas por Mäkinen (2002) en Betula pendula para rodales de 400, 1.600 y 5.000 árboles por hectárea y puede explicarse por la alta densidad de estas plantaciones que ejercen un alto control de la vitalidad de las ramas basales (Kozlowski y Pallardy 1997, Lowell et al. 2014). La longevidad de las ramas encontradas en este estudio está en directa relación con la edad del árbol como lo señala Uri (1998). N. obliqua en este estudio presenta edad de ramas muy inferiores a lo encontrado por Ishii y Wilson (2001) en P. menziesii. La vitalidad de las ramas es una característica específica. La correlación negativa entre edad de las ramas con la altura relativa de inserción, que se determinó en este estudio, está de acuerdo con lo reportado por estos mismos autores y es consistente con la positiva correlación encontrada en el presente estudio con el diámetro del fuste donde la rama se inserta.
El aumento del ángulo de inserción con la altura relativa de la rama constituye una evidencia del cambio ontogénico que N. obliqua presenta siendo más jerárquico a medida que aumenta con la edad. Esta evidencia está de acuerdo con lo señalado por Puntieri et al. (2013) y reafirmaría el carácter de intolerante de la especie y, por lo tanto, el efecto que la luminosidad tiene sobre el tamaño y ángulo de inserción de la ramas.
CONCLUSIONES
Del estudio realizado sobre la rama viva de mayor diámetro de cada decil de altura de 59 árboles se concluye que:
Dentro del árbol: el diámetro de la rama aumenta a medida que baja en altura relativa, que aumenta el diámetro fustal donde se inserta y que disminuye el ángulo de inserción. El menor ángulo de inserción de las ramas se encuentra en secciones fustales de mayor diámetro, en ramas más viejas y en posiciones inferiores de altura relativa. Las ramas más viejas tienen un diámetro mayor, se presentan en secciones fustales de mayor diámetro y en posiciones de altura relativa inferiores. Todas estas relaciones son significativas.
Entre árboles: el diámetro de la rama es mayor en árboles de mayor DAP, de mayor longitud fustal y de mayor edad. Las ramas de menor ángulo de inserción se ubican en árboles de DAP y longitud fustal mayor. Las ramas más viejas se presentan en árboles de mayor DAP y de mayor edad. Estas relaciones son significativas.
AGRADECIMIENTOS
Al Fondo de Investigación del Bosque Nativo que financió la toma de datos.
REFERENCIAS BIBLIOGRÁFICAS
CHANGSHENG, L.S., JIANFENG, X., YONGFANG, C., FRANCIS, H. and FRANCOIS, H. 1998. Crown morphology of Norway spruce from usual tree measurements. Journal of Forestry Research,1998, 9(1): 8-12. ISSN 0022-1201.
COLIN, F. and HOULLIER, F. 1992. Branchiness of Norway spruce in northeastern France: predicting the main crown characteristics from usual tree measurements. Annales des Sciences Forestières 49(5): 511-538.
Corvalán, P. 2015. Diagrama de manejo de la densidad de rodal para el control del tamaño de ramas basales en bosques septentrionales altoandinos dominados por roble en la región del Maule. Serie biomasa. Santiago, Chile. Facultad de Ciencias Forestales y de la Conservación de la Naturaleza, Universidad de Chile. 121 p. ISBN: 978-956-358-953-5.
Donoso, C. 1993. Bosques templados de Chile y Argentina. Variación, estructura y dinámica. Ecología Forestal. Santiago, Chile. Editorial Universitaria. 483 p. ISBN 956-11-0926-3.
Ford, E.D. 1985. Branching, crown structure and the control of timber production. In Cannell MGR, JE Jackson eds. Attributes of trees as crop plants. Huntingdon, UK. Abbotts Ripton, Institute of Terrestrial Ecology. p. 228-252.
GAJARDO, R. 1994. La vegetación natural de Chile. Clasificación y distribución geográfica. Santiago, Chile. Editorial Universitaria. 165 p.
GARBER, S. and MAGUIRE, D. 2005. Vertical trends in maximum branch diameter in two mixed-species spacing trials in the central Oregon Cascades. Canadian Journal of Forest Research 35(2): 295-307. DOI: 10.1139/X04-164.
GROOT, A. and SCHNEIDER, R. 2011. Predicting maximum branch diameter from crown dimensions, stand characteristics and tree species. The Forestry Chronicle 87(4): 542-551.
GROSSE, H. y Quiroz, I. 1999. Silvicultura de los bosques de segundo crecimiento de roble, raulí y coigüe en la región centro-sur de Chile. In Donoso C, A Lara eds. Silvicultura de los Bosques Nativos de Chile. Santiago, Chile. Editorial Universitaria. p. 95-128. ISBN 956-11-1425-9.
GSPALTL, M., BAUERLE, W., BINKLEY, D., STERBA, H. 2013. Leaf area and light use efficiency patterns of Norway spruce under different thinning regimes and age classes. Forest Ecology and Management 288: 49-59. DOI :10.1016/j.foreco.2011.11.044.
ISHII, H. and WILSON, M.E. 2001. Crown structure of old-growth Douglas-fir in the western Cascade Range, Washington. Canadian Journal of Forest Research 31(7): 1250-1261. doi:10.1139/cjfr-31-7-1250.
KANTOLA, A. and MÄKELÄ, A. 2004. Crown development in Norway spruce [Picea abies (L.) Karst.]. Trees 18(4): 408-421. DOI 10.1007/s00468-004-0319-x.
KOZLOWSKI, T.T. and PALLARDY, S.G. 1997. Growth control in woody plants. San Diego, USA. Academic Press. 641 p. ISBN 0-12-424210-3.
LARSON, P.R., KRETSCHMANN, D.E., CLARK, A.I. and ISEBRANDS, J.G. 2001. Formation and properties of juvenile wood in southern pines: a synopsis. USDA Forest Service General Technical Report FPL-GTR-129. Madison, USA. Department of Agriculture, Forest Service, Forest Products Laboratory. 42 p.
LOWELL, E.C., MAGUIRE, D.A., BRIGGS, D.G., TURNBLOM, E.C., JAYAWICKRAMA, K.J. and BRYCE, J. 2014. Effects of silviculture and genetics on branch/knot attributes of coastal Pacific Northwest Douglas-fir and implications for wood quality- A Synthesis. Forests 5(7): 1717-1736. doi:10.3390/f5071717.
LUEBERT, F. y PLISCOFF, P. 2006. Sinopsis bioclimática y vegetacional de Chile. Editorial Universitaria.
MAGUIRE, D.A., KERSHAW, J.A., HANN, D.W. 1991. Predicting the effects of silvicultural regime on branch size and crown wood core in Douglas-fir. Forest Science 37(5): 1409-1428.
MÄKINEN, H. and COLIN, F. 1999. Predicting the number, death, and self-pruning of branches in Scots pine. Canadian Journal of Forest Research 29(8): 1225-1236. doi.org/10.1139/x99-065.
MÄKINEN, H. 2002. Effect of stand density on the branch development of silver birch (Betula pendula Roth) in central Finland. Trees 16(4-5): 346-353. DOI 10.1007/s00468-002-0162-x.
NELSON, A.S., WEISKITTEL, A.R. and WAGNER, R.G. 2014. Development of branch, crown, and vertical distribution leaf area models for contrasting hardwood species in Maine, USA. Trees 28(1): 17-30. DOI 10.1007/s00468-013-0926-5.
PUNTIERI, J., TORRES, C. and GHIRARDI, S. 2013. Crecimiento y ramificación de Nothofagus alpina y Nothofagus obliqua (Nothofagaceae) bajo diferentes condiciones lumínicas. Growth and branching in Nothofagus alpina and Nothofagus obliqua (Nothofagaceae) under different light conditions. Boletín de la Sociedad Argentina de Botánica 48(2): 277-293. ISSN 0373-580X.
READ, J. and HILL, R. 1985. Photosynthetic responses to light of Australian and Chilean species of Nothofagus and their relevance to the rainforest dynamics. The New Phytologist 101(4): 731-742.
ROEH, R.L. and MAGUIRE, D.A. 1997. Crown profile models based on branch attributes in coastal Douglas-fir. Forest Ecology and Management 96(1): 77-100.
RUSSELL, M.B. and WEISKITTEL, A.R. 2011. Maximum and largest crown width equations for 15 tree species in Maine. Northern Journal of Applied Forestry 28(2): 84-91.
SONDERMAN, D.L. 1984. Quality response of even-aged 80-year-old white oak trees after thinning. Broomall, PA, USA. USDA, Forest Service, Northeastern Forest Experiment Station. 6 p.
TRINCADO, G. and BURKHART, H.E. 2008. A model of knot shape and volume in loblolly pine trees. Wood and Fiber Science 40(4): 634-646.
URI, V. 1998. The effect of thinning on the branch diameter increment in pine stands. Baltic Forestry 4 (1):17-23. ISSN 1392-1 355.