Evidencias de facilitación entre plantas en las arenas cuarcíticas de San Ubaldo
Revista Cubana de Ciencias Forestales. 2017, 5(1):3-14![]()
Evidencias de facilitación entre plantas en las arenas cuarcíticas de San Ubaldo
Evidences of facilitation among plants in the quarzitic sands of San Ubaldo
Enrique González Pendás1, Vidal Pérez Hernández2, Jorge Ferro Díaz3, Roberto J. Gómez Marín4
1Licenciado en Biología, Máster en Ciencias Forestales. Jardín Botánico de Pinar del Río, Km 2 ½ camino al Hoyo Guamá.
Correo electrónico: enrique@jbpr.vega.inf.cu
2Licenciado en Biologia. Doctor en Ciencias. Centro de Investigaciones Ecovida.
Correo electrónico: vidal@jbpr.vega.inf.cu
3Licenciado en Geografía. Doctor en Ciencias Forestales.
Correo
electrónico: ferro@ecovida.vega.inf.cu
4Ingeniero Forestal. Máster en Ciencias Forestales. Profesor de Manejo de Cuencas. Universidad de Pinar del Río «Hermanos Saíz Montes de Oca».
Correo
electrónico: rgomez@upr.edu.cu
Recibido: 6 de octubre de 2016.
Aprobado: 6 de marzo de 2017.
RESUMEN
La investigación se realiza en el área protegida de San Ubaldo, hacia el interior de un pinar abierto, con palmas en las llanuras secas sobre arenas cuarcíticas, pertenecientes a la reserva Florística Manejada Sabanalamar-San Ubaldo. El objetivo de esta investigación es determinar las evidencias de facilitación planta-planta y su relación con la estructura y composición de las comunidades vegetales presentes en estos peculiares ecosistemas. Para su estudio se trazan seis transeptos lineales paralelos de 100 m. de largo, en los que se realiza un levantamiento general de la flora. Se definieron cuáles especies se desempeñan como nodrizas y facilitadas, a partir de un estudio del número de individuos por especie, su frecuencia y abundancia, determinando las que más reclutan otras especies (mejores nodrizas). Se ofrece un listado florístico actualizado y se comprueba que la mediana de la riqueza específica de especies bajo las nodrizas es significativamente superior a la de los espacios abiertos; se demuestra que las especies, que conviven debajo de las nodrizas, no están filogenéticamente emparentadas; esto pudiera indicar las relaciones de facilitación y la importancia del efecto nodriza en interacciones típicas de ambientes extremos.
Palabras clave: facilitación, nodrizas, arenas cuarcíticas.
ABSTRACT
The study was carried out in the protected area of San Ubaldo, toward the interior of an open Pine grove with Palm trees in the dry plains over quartzite sands , belonging to the Floristic Managed Reservation Sabanalamar-San Ubaldo, the objective was to determine evidences of facilitation plant-plant and its relationship with the structure and composition of the present plants communities in these peculiar ecosystems; for its study, six lineal parallel transepts of 100 m is traced, in those it is carried out a general rising of the flora, They were defined which species act as wet nurses and facilitated, from a study of the number of individuals per species, their frequency and abundance, determining the most recruit other species (best wet nurses). A floristic updated list is available; it is found that the median wealth of specific species under suckler is significantly higher than the open spaces and it shows that the species that live beneath the wet nurses are not phylogenetically related. This is an evidence of relationships of facilitation among plants, as well as the importance of the effect nurse in typical interactions on extreme environments.
Key words: Facilitation, nurse plants, quartzite sands.
INTRODUCCIÓN
El área de estudio se encuentra en la Reserva Florística Manejada Sabanalamar-San Ubaldo, que se localiza en el municipio Güane, Pinar del Río. De acuerdo con Novo & Luis (1989), esta zona se caracteriza por presentar drásticas variaciones estacionales, elevadas temperaturas y un nivel de evapotranspiración muy alto, influenciado, según Borhidi (1996), por la presencia de suelos areno cuarcíticos casi puros, muy ácidos y pobres en nutrientes.
Esta alternancia de etapas secas y húmedas en la llanura favorece la existencia de condiciones extremas para la biota; por una parte, las inundaciones durante el periodo húmedo mientras durante el periodo seco la friabilidad de las arenas hace que el agua se escurra con rapidez. De esta manera, la mayoría de las especies son anuales o bienales y se convierten estos ecosistemas extremos en relictos de alto endemismo vegetal, según Novo & al. (1984).
De acuerdo con Ricardo & al. (2009), la vegetación de este tipo de zonas está representada por pinares abiertos y sabanas seminaturales con palmas, además de diversos cuerpos de agua con su propia vegetación asociada. En esta área de estudio, en el pinar abierto con palmas, predominan los arbustos; también se pueden encontrar sinusias herbáceas en dependencia del nivel de antropización; en lugares mejor conservados solo se observan «arenasos abiertos» entre los grupos aislados de arbustos y pinos.
Teniendo en cuenta los criterios de Oviedo &al. (2014), las interacciones ecológicas, intra e ínterespecífica, se consideran esenciales para entender la evolución biológica y el mantenimiento de la diversidad en la naturaleza.
Por otra parte, Tirado & Pugnaire (2005) y Verdú & Valiente-Banuet (2008) plantean que la facilitación ha sido expuesta repetidamente como un importante mecanismo de estructuración, en comunidades de plantas, bajo condiciones estresantes, por lo que se decide estudiar este caso de ambientes extremos, para determinar las evidencias de facilitación entre plantas y su relación con la estructura y composición de las comunidades vegetales presentes en estos peculiares ecosistemas, argumentos importantes a tener en cuenta a la hora de tratar de restaurarlos, sabiendo que es muy complicado el cultivo exsitu de la flora presente en los mismos.
MATERIALES Y METODOS
En la zona de estudio, caracterizada por pinares abiertos con palmas, se obtiene producto del registro realizado, un listado florístico actualizado (Ver tabla 1.), de acuerdo con los cambios taxonómicos aprobados por el APG IV. (Angiosperm Phylogeny Group), además, según la metodología de trabajo empleada por Oviedo &al. (2014), en la cual se plantea trazar seis transeptos paralelos de 100 m. cada uno; estimar la cobertura vegetal por el método de la línea transepta (Canfield, 1941); definir que cada planta determinada como nodriza representa un parche, así como los grupos compactos de plantas de la misma especie; asumir que funcionan como una, además de registrar los espacios abiertos.
Se tiene en cuenta, como patrón para la comparación entre parches, multiespecies con respecto a los espacios abiertos: el rango de riqueza específica, la mediana de la riqueza específica, el rango de abundancia y la mediana de la abundancia, los cuales se muestran en una tabla comparativa (Tabla 1).
Las distancias filogenéticas se calculan, a partir de un mega árbol, con la ayuda del programa Phylomatic, versión 4.2 Phylocom Webb & al. (2008). El árbol filogenético de la comunidad se obtiene al emparejar los nombres de las familias de plantas presentes en el sitio de estudio, con los incluidos en un mega árbol de angiospermas, que se utiliza en Phylomatic (R20120829. new), basado en el trabajo de Stevens (2001).
Las estimaciones de las edades en las familias de plantas se utilizan para datar el árbol, según Wikstrom & al. (2001), y se aplica el algoritmo de Bladj, que distribuye nodos sin fechas uniformes entre los nodos de edades conocidas, según Webb & al. (2008).
ANÁLISIS Y DISCUSIÓN DE LOS RESULTADOS
Como resultado del registro realizado se obtienen los siguientes datos:
Tabla 1. Listado florístico actualizado de la zona de estudio.
ESPECIES |
FAMILIAS |
ABUNDANCIA |
1. Anacardium occidentale L. |
Anacardiaceae |
1 |
2. Xylopia aromatica (Lam.) Mart. |
Annonaceae |
1 |
3. Cynanchum sp. |
Apocynaceae |
40 |
4. Cynanchum savannarum Alain. |
Apocynaceae |
6 |
5. Mesechites rosea (A.DC.) Miers. |
Apocynaceae |
1 |
6. Ilex cassineL. |
Aquifoliaceae |
2 |
7. Colpothrinax wrightii Griseb. &H.Wendl. |
Arecaceae |
75 |
8. Acoelorraphe wrightii (Griseb. &H.Wendl.) |
Arecaceae |
91 |
9. Coccothrinax miraguama subsp. |
Arecaceae |
5 |
10. Eupatorium capilifolium(Lam.) Small. |
Asteraceae |
1 |
11. Aster grsisebachii Britton. |
Asteraceae |
31 |
12. Eupatorium odoratum L. |
Asteraceae |
1 |
13. Emilia sagittata(Vahl) DC. |
Asteraceae |
4 |
14. Tabebuia lepidophylla(A.Rich.) Greenm. |
Bignoniaceae |
13 |
15. Blechnum serrulatum L.C. Rich. |
Blechnaceae |
258 |
16. Burmannia capitata (Walter) Mart. |
Burmanniaceae |
3 |
17. Opuntia stricta Haw. |
Cactaceae |
14 |
18. Harrisia eriophora(Pfeiff.) Britton. |
Cactaceae |
6 |
19. Chrisobalanus icaco L. |
Chrisobalanaceae |
24 |
20. Lechea cubenisL egg. |
Cistaceae |
11 |
21. Rhynchospora fascicularis (Michx.) Vahl. |
Cyperaceae |
293 |
22. Cyperus sp. |
Cyperaceae |
1 |
23. Eleocharis minima Kunth. |
Cyperaceae |
8 |
24. Drosera capillaris Poir. |
Droseraceae |
72 |
25. Kalmia ericoides C.Wright ex Griseb. |
Ericaceae |
15 |
26. Paepalanthus sp. |
Eriocaulaceae |
10 |
27. Syngonanthus androsaceus(Griseb.) |
Eriocaulaceae |
52 |
28. Eriocaulonsigmoideum C.Wright. |
Eriocaulaceae |
146 |
29. Paepalanthuslamarckii Kunth. |
Eriocaulaceae |
70 |
30. Croton craspedotrichus Griseb. |
Euphorbiaceae |
3 |
31. Croton cerinus Müll.Arg. |
Euphorbiaceae |
302 |
32. Chamaesyce pinariona (Urb.) Alain. |
Euphorbiaceae |
10 |
33. Clitoria sp. |
Fabaceae |
7 |
34. Chamaecrista diphylla (L.) Greene. |
Fabaceae |
47 |
35. Brya ebenus L. |
Fabaceae |
106 |
36. Dichrostachys cinerea (L.) Wight &Arn. |
Fabaceae |
252 |
37. Indigofera miniata Ortega. |
Fabaceae |
37 |
38. Cleome guianensis Aubl. |
Fabaceae |
21 |
39. Chamaecrista pilosa (L.) Greene |
Fabaceae |
304 |
40. Clitoria laurifolia Poir. |
Fabaceae |
12 |
41. Chamaecrista sp. |
Fabaceae |
10 |
42. Aeschynomene viscidula Michx. |
Fabaceae |
14 |
43. Quercus cubana A. Rich. |
Fagaceae |
1 |
44. Lachnanthes caroliniana (Lam.) Dandy |
Haemodoraceae |
38 |
45. Hypericum stiphelioides A.Rich. |
Hypericaceae |
297 |
46. Hypericum nitidum Lam. |
Hypericaceae |
65 |
47. Cassytha filiformis L. |
Lauraceae |
4 |
48. Pinguicula albida C.Wright ex Griseb. |
Lentibulariaceae |
3 |
49. Utricularia juncea Vahl. |
Lentibulariaceae |
22 |
50. Polypremum procumbens L. |
Loganiaceae |
112 |
51. Lycopodium carolineanum L. |
Lycopodiaceae |
47 |
52. Byrsonima crassifolia (L.) |
Malpighiaceae |
51 |
53. Byrsonima coccolobaefolia |
Malpighiaceae |
2 |
54. Malpighia horrida Small. |
Malpighiaceae |
4 |
55. Byrsonima wrightiana Urb. &Nied. |
Malpighiaceae |
2 |
56. Sida brittonii León. |
Malvaceae |
5 |
57. Waltheria indica A. Rodr. |
Malvaceae |
26 |
58. Melochia sp. |
Malvaceae |
2 |
59. Melochia villosa (Mill.) Fawc. |
Malvaceae |
27 |
60. Peltaea speciosa (Kunth) Standley |
Malvaceae |
2 |
61. Hibiscus costatus A.Rich. |
Malvaceae |
2 |
62. Urena lobata L. |
Malvaceae |
1 |
63. Pachyanthus wrightii Griseb. |
Melastomataceae |
11 |
64. Chaetolepis cubensis (A.Rich.) Triana. |
Melastomataceae |
45 |
65. Acisanthera quadrata Juss. |
Melastomataceae |
5 |
66. Cecropia schreberiana Miq. |
Moraceae |
3 |
67. Myrica cerífera L. |
Myricaceae |
35 |
68. Eucalyptus citriodora Hook. |
Myrtaceae |
4 |
69. Eugenia punicifolia(Humb., Bonpl. & Kunth) DC. |
Myrtaceae |
6 |
70. Sauvagesiabrowneii Planch. |
Ochnaceae |
93 |
71. Platanthera replicata (Rich.) Ackerman. |
Orquidaceae |
4 |
72. Phyllantus equinospermus C.Wright ex Sauvalle. |
Phyllantaceae |
21 |
73. Pinus caribaea var. CaribaeaMorelet. |
Pinaceae |
13 |
74. Pinus tropicalis Morelet. |
Pinaceae |
39 |
75. Paspalum notatum Flüggé. |
Poaceae |
430 |
76. Panicum erectifolium Nash. |
Poaceae |
124 |
77. Mitracarpuss caberullus Urb. |
Rubiaceae |
8 |
78. Hedyotis callitrichoides (Griseb.) W.H.Lewis |
Rubiaceae |
13 |
79. Diodia teres Walter. |
Rubiaceae |
22 |
80. Smilax havanensis Jacq. |
Smilacaceae |
14 |
81. Waltheria arenícola A.Rodr. |
Sterculiaceae |
24 |
82. Thelypteris interrupta (Willd.) K.Iwats. |
Thelypteridaceae |
2 |
83. Lantana strigosa (Griseb.) Urb. |
Verbenaceae |
1 |
84. Xyris jupicai Rich. |
Xyridaceae |
33 |
85. Xyris navicularis Griseb. |
Xyridaceae |
31 |
86. Xyris grandiceps Griseb. |
Xyridaceae |
1 |
87. Xyrisbissei Urquiolaetkral. |
Xyridaceae |
8 |
88. Xyris brevifolia Michx. |
Xyridaceae |
39 |
89. Xyris elliotii Chapm. |
Xyridaceae |
22 |
- Especie de mayor abundancia: Paspalum notatum.
- Nodriza con más altos valores medios de la riqueza específica debajo (mejor reclutadora): Byrsonima crassifolia (10.4). (Figura 1.)
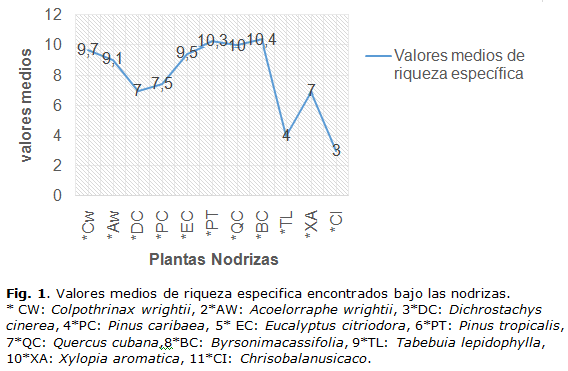
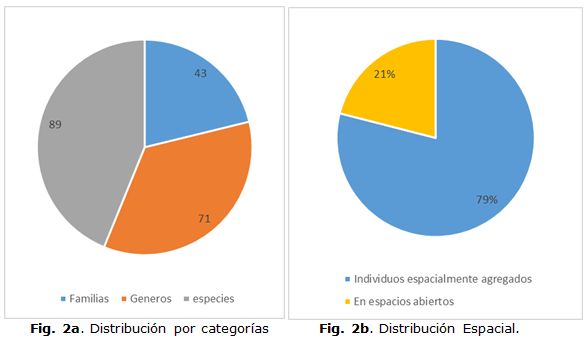
Se registraron 5509 individuos, pertenecientes a 89 especies, que se agrupan en 71 géneros y 43 familias (Figura 2a.). Al analizar la proporcionalidad entre las categorías taxonómicas, en la evaluación de la estructura filogenética realizada a la diversidad de esta comunidad, se determina que no existe una clara estructura anidada entre ellas, como se aprecia en otras comunidades en ambientes menos extremos.
Es importante tener en cuenta que, a medida que aparezcan nuevas especies durante el muestreo, se corresponden con la aparición de nuevos géneros y familias, lo que favorece a una mayor diversidad filogenético en concordancia con lo planteado por Callaway (2007), que asume este comportamiento como típico, en comunidades biológicas que se desarrollan en ecosistemas de elevado estrés, donde los efectos de la facilitación impulsan la dinámica funcional y anteceden a otros procesos como los de competencia.
El 78,9% de los individuos (4347) viven espacialmente agregados con otras especies, formando un total de 59 parches multiespecies. Por otra parte, 1162 individuos (21,1%) viven en los 50 espacios abiertos registrados (Figura 2b.).
De acuerdo con la cobertura que presentan las especies y siguiendo los criterios de Oviedo & al. (2014), se registraron 11 especies que actúan como nodrizas y, debajo de ellas, se observan 78 especies que se consideran como facilitadas, una cifra mayor que la de estudios realizados en el mediterráneo español que presentaba el segundo caso con mayor cantidad de nodrizas (11) y especies facilitadas (45), solo superada, hasta la fecha, por los mecanismos de facilitación descritos en el desierto de Sonora, con (29) nodrizas y (177) especies facilitadas, según Flores & Jurado (2003).
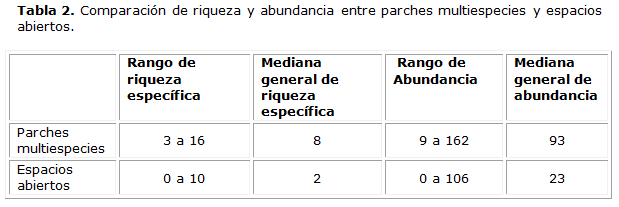
El valor para la mediana de la riqueza específica debajo de las nodrizas resulta de ocho (8), mientras que en los espacios vacíos es dos (2). La alta concentración de individuos, con distribución espacial agrupada alrededor de otras especies, se considera como una muestra de que la estructura y composición en esta comunidad están basadas en la existencia de facilitación entre las plantas, beneficiadas por el aprovechamiento de la cobertura y, con ello, las facilidades ecológicas inducidas.
En la tabla 2, se muestra el valor más alto en cuanto a riqueza específica (16) de las mejores nodrizas, muy superior al valor de (10), registrado en los espacios vacíos. Esto demuestra que las especies vegetales conviven más agrupadas que aisladas, una evidencia de facilitación como interacción ensambladora de comunidades en ambientes extremos, compartiendo los criterios que aportan Bertness & Callaway (1994); Broker & Callaghan (1998); Pugnaire & Luque (2001) y Bruno & al. (2003).
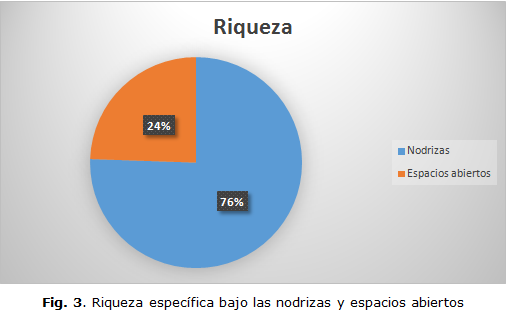
Una fuerte evidencia de la existencia y papel de la facilitación como proceso ecológico destacado se puede apreciar en la distinción de distribución de la riqueza específica total encontrada, tanto en parches de vegetación (bajo nodrizas), como en espacios abiertos, sin presencia de facilitación, lo cual se muestra en el esquema de distribución porcentual de esas riquezas (Figura 3).
En el árbol de la comunidad (Ver figura 4)se puede apreciar una alta filo
diversidad típica de casos de facilitación, en
donde esta tiende a unir a parientes lejanos, ya
que tienen nichos de regeneración complementarios (Verdú & Valiente-Banuet. 2008);
en tanto, el árbol de Byrsonimacrassifolia, el cual resulta la nodriza mejor reclutadora,
deja ver que la diversidad filogenética del parche aumenta con la riqueza de especies, lo
que concuerda con lo expuesto por Callaway (2007) acerca de que interacciones indirectas,
a través de terceras especies de plantas, suelen aliviar la competición por
parejas, además, es necesario apuntar que plantas como el caso de Byrsonimacrassifolia han demostrado ser buenas enriquecedoras del suelo (Kellman 1979): una de
las características que debe reunir una buena nodriza (Flores & Jurado. 2003).
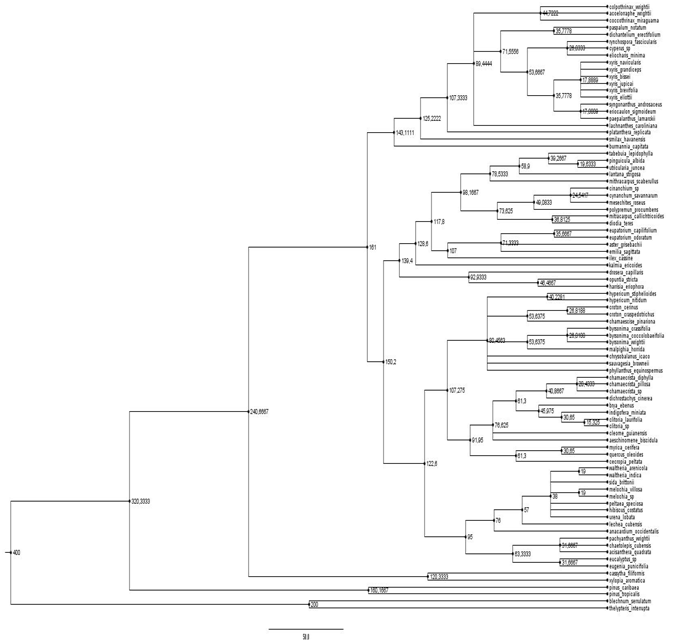
Fig. 4. Árbol filogenético de la comunidad de plantas estudiadas.
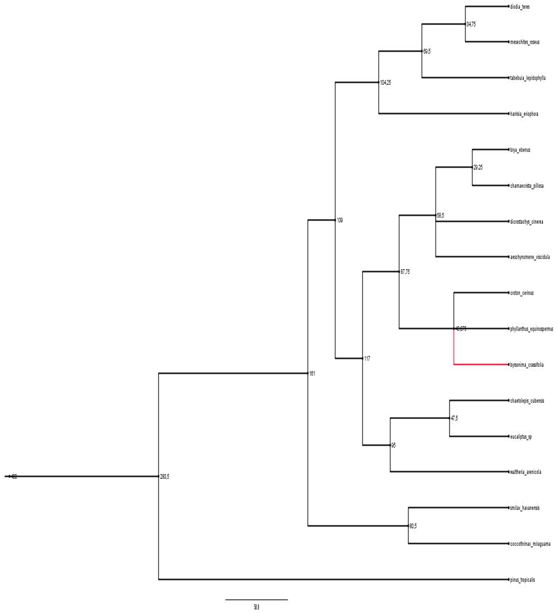
Fig. 5. Árbol filogenético donde Byrsonima crassifolia actúa como nodriza.
Es la facilitación, y no la competencia, el proceso ecológico que establece el ensamblaje de esta comunidad vegetal del área de San Ubaldo, en la cual domina el efecto nodriza como motor de ensamblaje y, a su vez, como mecanismo de preservación de la diversidad vegetal, al favorecer el establecimiento de plántulas de diferentes especies bajo condiciones más favorables, como ya ha sido planteado anteriormente para ecosistemas de condiciones extremas, tal y como es el caso de nuestra área de estudio.
Resalta el caso de Byrsonima crassifolia como una buena planta nodriza, aspecto a considerar para su uso en planes de restauración o reforestación de la zona.
REFERENCIAS BIBLIOGRÁFICAS
[1] BERTNESS, M. D. y CALLAWAY, R. M. Positive interactions in communities. Trends in Ecology & Evolutión [En línea]. 1994, mayo, 9(5), 191-193. ISSN: 0169-5347. Disponible en: http://www.sciencedirect.com/science/article/pii/0169534794900884
[2] BORHIDI A. L. Phytogeography and vegetation ecology of Cuba. Budapest, Hungary: Akademia Kiado, 1991. ISBN 9630552957
[3] BROOKER, R. W. y CALLAGHAN, T. V. The balance between positive and negative plant interactions and its relationship to environmental gradients: A Model. Oikos [En línea] .1998, febrero, 81(1). 196-207. doi: 10.2307/3546481. Disponible en: http://www.jstor.org/stable/3546481?seq=1#page_scan_tab_contents
[4] BRUNO, J. F, STACHOWICZ, J. J. y BERTNESS, M. D. Inclusion of facilitation into ecological theory. Trends in Ecology & Evolutión [En línea]. 2003, marzo, 18(3), 119-125. ISSN: 0169-5347. Disponible en: http://www.sciencedirect.com/science/article/pii/S0169534702000459
[5] CALLAWAY, R. M. Positive interactions and interdependence in plant communities. Montana: Springer Netherlands, 2007. ISBN: 978-1-4020-6223-0
[6] CANFIELD R. H., Application of the line intercepts method in sampling range vegetation. Journal of Forestry [En línea]. 1941, abril, 39 (4). 388-394. ISSN 0022-1201 .Disponible en: http://www.ingentaconnect.com/content/saf/jof/1941/00000039/00000004/art00012
[7] FLORES, J. y JURADO, E. Are nurse-protégé interactions more common among plantsfrom arid environments? Journal of Vegetation Science [En línea]. 2003, diciembre, 14(6). 911-916. ISSN1654-1103. Disponible en: http://www.bioone.org/doi/abs/10.1658/1100-9233%282003%29014[0911:ANIMCA]2.0.CO%3B2
[8] KELLMAN, M. Soil enrichment by Netropical savanna trees, Journal of Ecology [En línea]. Julio, 1979, 67(2). 565-577. Disponible en: http://www.jstor.org/stable/2259112#?seq=1#page_scan_tab_contents
[9] NOVO R, LUÍS M., URQUIOLA A. y VEGA E.: Análisis paleogeográfico de las Llanuras arenosas de Pinar del Río. Instituto Superior Pedagógico de Pinar del Río: Departamento de Geografía-Centro de Documentación., 1984.
[10] OVIEDO PRIETO R., FAIFE CABRERA M., NOA MONZÓN A., ARROYO J., VALIENTE-BANUET A. Facilitation allows plant coexistence in Cuban serpentine soils. Plant Biology, 2014, Julio, 16(4). 711716. ISSN 1435-8603. Disponible en: http://onlinelibrary.wiley.com/doi/10.1111/plb.12116/full
[11] PUGNAIRE, F. I. & LUQUE, M. T. Changes in plant interactions along a gradient of environmental stress./Oikos, 2001, abril, 93(1). 42-49. DOI: 10.1034/j.1600-0706.2001.930104.x. Disponible en: http://onlinelibrary.wiley.com/doi/10.1034/j.1600-0706.2001.930104.x/full
[12] RAMÍREZ CONTRERAS, A. y RODRÍGUEZ TREJO, D. A. Plantas nodriza en la reforestación con PinushartwegiiLindl. Revista Chapingo ,2009, 15 (1). 43-48. ISSN: 0186-3231
[13] RICARDO NÁPOLES N.E., et al. Tipos y características de las formaciones vegetales de Cuba. Acta Botánica Cubana [En línea]. 2009. Disponible en: http://repositorio.geotech.cu/xmlui/handle/1234/660
[14] STEVENS, P.F. Angiosperm Phylogeny Website. Version13. [En línea]. Missouri Botanical Garden: University of Missouri, St Louis. [consultado 19 febrero 2013] Disponible en: http://www.mobot.org/MOBOT/research/APweb/
[15] TIRADO R. & PUGNAIRE F.I. Community estructure and positive interactions in constraining environments. Oikos [En línea]. 2005, diciembre, 111(3). 437-444. DOI: 10.1111/j.1600-0706.2005.14094.x. Disponible en: http://onlinelibrary.wiley.com/doi/10.1111/j.1600-0706.2005.14094.x/full
[16] VALIENTE-BANUET A., et al. Modern Quaternary plant lineages promote diversity through facilitation of ancient Tertiary lineages. Proceedings of the National Academy of Sciences USA [En línea]. 2006, 103(45). 1681216817. SSN 1091-6490. DOIi: 10.1073/pnas.0604933103. Disponible en: http://www.pnas.org/content/103/45/16812.short
[17] VALIENTE-BANUET A. y VERDÚ M. Facilitation can increase the phylogenetic diversity of plant communities. Ecology Letters [En línea]. 2007, noviembre, 10(11). 10291036. DOI: 10.1111/j.1461-0248.2007.01100.x. Disponible en: http://onlinelibrary.wiley.com/doi/10.1111/j.1461-0248.2007.01100.x/full
[18] VERDU M., VALIENTE-BANUET A. The nested assembly of plant facilitation networks prevents species extinctions. The American Naturalist [En línea]. 2008, diciembre, 172(6). 751760. DOI: 10.1086/593003. Disponible en: http://www.journals.uchicago.edu/doi/abs/10.1086/593003
[19] WEBB C.O., ACKERLY D.D. y KEMBEL S.W. Phylocom: software for the analysis of phylogenetic community structure and character evolution. Bioinformatics Applications Note [En línea]. 2008, agosto, 24(18). 20982100. doi:10.1093/bioinformatics/btn358. Disponible en: http://phylodiversity.net/cwebb/pubs/webb2008_phylocom.pdf
[20] WIKSTROM N., SAVOLAINEN V., CHASE M.W. Evolution of the angiosperms: calibrating the family tree. The Royal Society [En línea]. 2001, noviembre, 268(1482). 22112220. DOI:10.1098/rspb.2001.1782. Disponible en: http://rspb.royalsocietypublishing.org/content/royprsb/268/1482/2211.full.pdf